
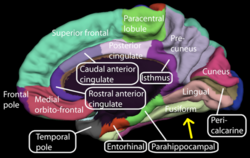
The cingulate cortex is a part of the brain situated in the medial aspect of the cerebral cortex. The cingulate cortex includes the entire cingulate gyrus, which lies immediately above the corpus callosum, and the continuation of this in the cingulate sulcus. The cingulate cortex is usually considered part of the limbic lobe.

A Brodmann area is a region of the cerebral cortex, in the human or other primate brain, defined by its cytoarchitecture, or histological structure and organization of cells. The concept was first introduced by the German anatomist Korbinian Brodmann in the early 20th century. Brodmann mapped the human brain based on the varied cellular structure across the cortex and identified 52 distinct regions, which he numbered 1 to 52. These regions, or Brodmann areas, correspond with diverse functions including sensation, motor control, and cognition.

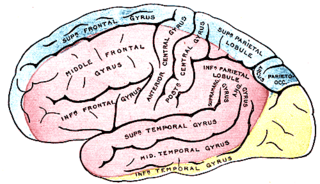
The temporal lobe is one of the four major lobes of the cerebral cortex in the brain of mammals. The temporal lobe is located beneath the lateral fissure on both cerebral hemispheres of the mammalian brain.
The frontal lobe is the largest of the four major lobes of the brain in mammals, and is located at the front of each cerebral hemisphere. It is parted from the parietal lobe by a groove between tissues called the central sulcus and from the temporal lobe by a deeper groove called the lateral sulcus. The most anterior rounded part of the frontal lobe is known as the frontal pole, one of the three poles of the cerebrum.

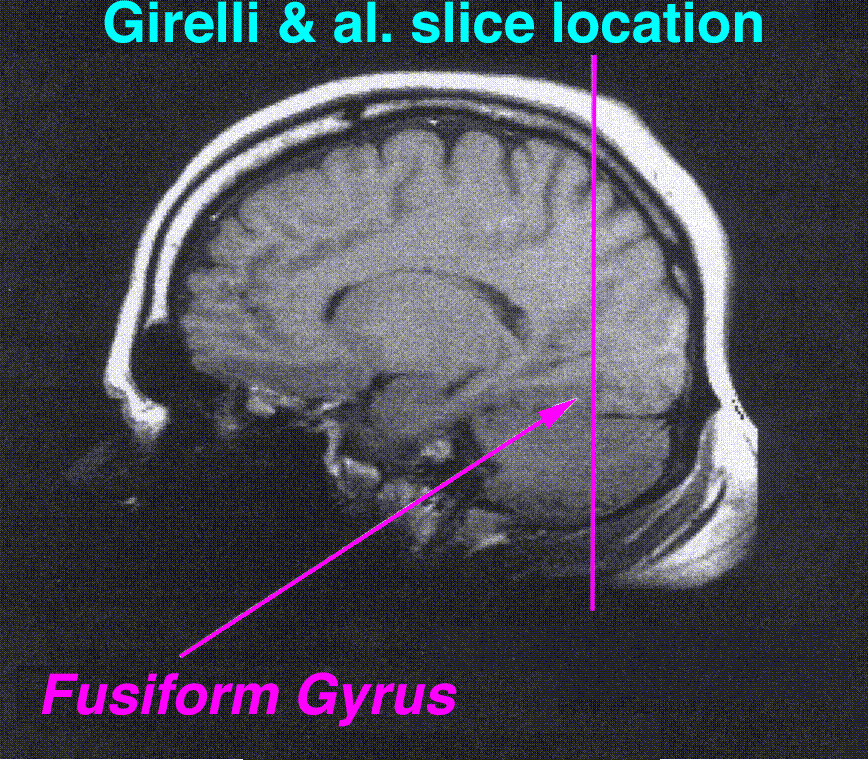
Brodmann area 37, or BA37, is part of the temporal cortex in the human brain. It contains the fusiform gyrus which in turn contains the fusiform face area, an area important for the recognition of faces.

In neuroanatomy, a gyrus is a ridge on the cerebral cortex. It is generally surrounded by one or more sulci. Gyri and sulci create the folded appearance of the brain in humans and other mammals.

The parahippocampal gyrus is a grey matter cortical region of the brain that surrounds the hippocampus and is part of the limbic system. The region plays an important role in memory encoding and retrieval. It has been involved in some cases of hippocampal sclerosis. Asymmetry has been observed in schizophrenia.

The lobes of the brain are the major identifiable zones of the human cerebral cortex, and they comprise the surface of each hemisphere of the cerebrum. The two hemispheres are roughly symmetrical in structure, and are connected by the corpus callosum. They traditionally have been divided into four lobes, but are today considered as having six lobes each. The lobes are large areas that are anatomically distinguishable, and are also functionally distinct to some degree. Each lobe of the brain has numerous ridges, or gyri, and furrows, the sulci that constitute further subzones of the cortex. The expression "lobes of the brain" usually refers only to those of the cerebrum, not to the distinct areas of the cerebellum.
The posterior cerebral artery (PCA) is one of a pair of cerebral arteries that supply oxygenated blood to the occipital lobe, part of the back of the human brain. The two arteries originate from the distal end of the basilar artery, where it bifurcates into the left and right posterior cerebral arteries. These anastomose with the middle cerebral arteries and internal carotid arteries via the posterior communicating arteries.

Brodmann area 35, together with Brodmann area 36, comprise the perirhinal cortex. They are cytoarchitecturally defined temporal regions of the cerebral cortex.

Middle temporal gyrus is a gyrus in the brain on the temporal lobe. It is located between the superior temporal gyrus and inferior temporal gyrus. It corresponds largely to Brodmann area 21.

The inferior temporal gyrus is one of three gyri of the temporal lobe and is located below the middle temporal gyrus, connected behind with the inferior occipital gyrus; it also extends around the infero-lateral border on to the inferior surface of the temporal lobe, where it is limited by the inferior sulcus. This region is one of the higher levels of the ventral stream of visual processing, associated with the representation of objects, places, faces, and colors. It may also be involved in face perception, and in the recognition of numbers and words.

The inferior parietal lobule lies below the horizontal portion of the intraparietal sulcus, and behind the lower part of the postcentral sulcus. Also known as Geschwind's territory after Norman Geschwind, an American neurologist, who in the early 1960s recognised its importance. It is a part of the parietal lobe.

The lingual gyrus, also known as the medialoccipitotemporal gyrus, is a brain structure that is linked to processing vision, especially related to letters. It is thought to also play a role in analysis of logical conditions and encoding visual memories. It is named after its shape, which is somewhat similar to a tongue. Contrary to the name, the region has little to do with speech.
In neuroanatomy, a sulcus is a depression or groove in the cerebral cortex. It surrounds a gyrus, creating the characteristic folded appearance of the brain in humans and other mammals. The larger sulci are usually called fissures.

The superior longitudinal fasciculus (SLF) is an association tract in the brain that is composed of three separate components. It is present in both hemispheres and can be found lateral to the centrum semiovale and connects the frontal, occipital, parietal, and temporal lobes. This bundle of tracts (fasciculus) passes from the frontal lobe through the operculum to the posterior end of the lateral sulcus where they either radiate to and synapse on neurons in the occipital lobe, or turn downward and forward around the putamen and then radiate to and synapse on neurons in anterior portions of the temporal lobe.
Visual object recognition refers to the ability to identify the objects in view based on visual input. One important signature of visual object recognition is "object invariance", or the ability to identify objects across changes in the detailed context in which objects are viewed, including changes in illumination, object pose, and background context.
The mid-fusiform sulcus is a shallow sulcus that divides the fusiform gyrus into lateral and medial partitions. Functionally, the MFS divides both large-scale functional maps and identifies fine-scale functional regions such as the anterior portion of the fusiform face area.

The occipital gyri (OcG) are three gyri in parallel, along the lateral portion of the occipital lobe, also referred to as a composite structure in the brain. The gyri are the superior occipital gyrus, the middle occipital gyrus, and the inferior occipital gyrus, and these are also known as the occipital face area. The superior and inferior occipital sulci separates the three occipital gyri.
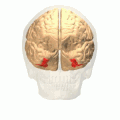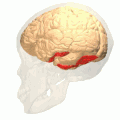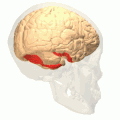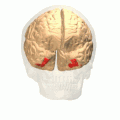
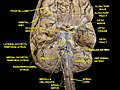
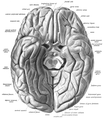
 Fusiform gyrus and adjacent gyri.fusiform gyrus; inferior temporal gyrus; parahippocampal gyrus; lingual gyrus; inferior occipital gyrus.
Fusiform gyrus and adjacent gyri.fusiform gyrus; inferior temporal gyrus; parahippocampal gyrus; lingual gyrus; inferior occipital gyrus.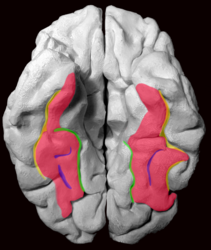 Fusiform gyrus and delimiting sulci.fusiform gyrus; occipitotemporal sulcus (OTS); collateral sulcus (CoS); mid-fusiform sulcus (MFS).
Fusiform gyrus and delimiting sulci.fusiform gyrus; occipitotemporal sulcus (OTS); collateral sulcus (CoS); mid-fusiform sulcus (MFS).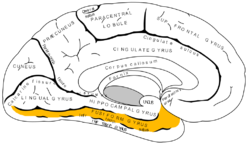






{kind=link}