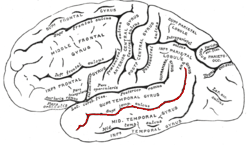
The auditory cortex takes part in the spectrotemporal, meaning involving time and frequency, analysis of the inputs passed on from the ear. The cortex then filters and passes on the information to the dual stream of speech processing.[5] The auditory cortex's function may help explain why particular brain damage leads to particular outcomes. For example, unilateral destruction, in a region of the auditory pathway above the cochlear nucleus, results in slight hearing loss, whereas bilateral destruction results in cortical deafness.
Structure
The auditory cortex was previously subdivided into primary (A1) and secondary (A2) projection areas and further association areas. The modern divisions of the auditory cortex are the core (which includes primary auditory cortex, A1), the belt (secondary auditory cortex, A2), and the parabelt (tertiary auditory cortex, A3). The belt is the area immediately surrounding the core; the parabelt is adjacent to the lateral side of the belt.[6]
Besides receiving input from the ears via lower parts of the auditory system, it also transmits signals back to these areas and is interconnected with other parts of the cerebral cortex. Within the core (A1), its structure preserves tonotopy, the orderly representation of frequency, due to its ability to map low to high frequencies corresponding to the apex and base, respectively, of the cochlea.
Like many areas in the neocortex, the functional properties of the adult primary auditory cortex (A1) are highly dependent on the sounds encountered early in life. This has been best studied using animal models, especially cats and rats. In the rat, exposure to a single frequency during postnatal day (P) 11 to 13 can cause a 2-fold expansion in the representation of that frequency in A1.[9] Importantly, the change is persistent, in that it lasts throughout the animal's life, and specific, in that the same exposure outside of that period causes no lasting change in the tonotopy of A1. Sexual dimorphism within the auditory cortex can be seen in humans between males in females through the planum temporale, encompassing Wernicke's region, for the planum temporale within males has been observed to have a larger planum temporale volume on average, reflecting previous studies discussing interactions between sex hormones and asymmetrical brain development.[10]
Function
As with other primary sensory cortical areas, auditory sensations reach perception only if received and processed by a cortical area. Evidence for this comes from lesion studies in human patients who have sustained damage to cortical areas through tumors or strokes,[11] or from animal experiments in which cortical areas were deactivated by surgical lesions or other methods.[12] Damage to the auditory cortex in humans leads to a loss of any awareness of sound, but an ability to react reflexively to sounds remains as there is a great deal of subcortical processing in the auditory brainstem and midbrain.[13][14][15]
Neurons in the auditory cortex are organized according to the frequency of sound to which they respond best. Neurons at one end of the auditory cortex respond best to low frequencies; neurons at the other respond best to high frequencies. There are multiple auditory areas (much like the multiple areas in the visual cortex), which can be distinguished anatomically and on the basis that they contain a complete "frequency map." The purpose of this frequency map (known as a tonotopic map) likely reflects the fact that the cochlea is arranged according to sound frequency. The auditory cortex is involved in tasks such as identifying and segregating "auditoryobjects" and identifying the location of a sound in space. For example, it has been shown that A1 encodes complex and abstract aspects of auditory stimuli without encoding their "raw" aspects like frequency content, presence of a distinct sound or its echoes.[16]
Human brain scans indicated that a peripheral part of this brain region is active when trying to identify musical pitch. Individual cells consistently get excited by sounds at specific frequencies, or multiples of that frequency.
The auditory cortex plays an important yet ambiguous role in hearing. When the auditory information passes into the cortex, the specifics of what exactly takes place are unclear. There is a large degree of individual variation in the auditory cortex, as noted by English biologist James Beament, who wrote, "The cortex is so complex that the most we may ever hope for is to understand it in principle, since the evidence we already have suggests that no two cortices work in precisely the same way."[17]
In the hearing process, multiple sounds are transduced simultaneously. The role of the auditory system is to decide which components form the sound link. Many have surmised that this linkage is based on the location of sounds. However, there are numerous distortions of sound when reflected off different media, which makes this thinking unlikely.[citation needed] The auditory cortex forms groupings based on fundamentals; in music, for example, this would include harmony, timing, and pitch.[18]
The auditory cortex is composed of fields that differ from each other in both structure and function.[19] The number of fields varies in different species, from as few as 2 in rodents to as many as 15 in the rhesus monkey. The number, location, and organization of fields in the human auditory cortex are not known at this time. What is known about the human auditory cortex comes from a base of knowledge gained from studies in mammals, including primates, used to interpret electrophysiological tests and functional imaging studies of the brain in humans.
When each instrument of a symphony orchestra or jazz band plays the same note, the quality of each sound is different, but the musician perceives each note as having the same pitch. The neurons of the auditory cortex of the brain are able to respond to pitch. Studies in the marmoset monkey have shown that pitch-selective neurons are located in a cortical region near the anterolateral border of the primary auditory cortex. This location of a pitch-selective area has also been identified in recent functional imaging studies in humans.[20][21]
The auditory cortex is the most highly organized processing unit of sound in the brain. This cortex area is the neural crux of hearing, and—in humans—language and music. The auditory cortex is divided into three separate parts: the primary, secondary, and tertiary auditory cortex. These structures are formed concentrically around one another, with the primary cortex in the middle and the tertiary cortex on the outside.
The primary auditory cortex is tonotopically organized, which means that neighboring cells in the cortex respond to neighboring frequencies.[23] Tonotopic mapping is preserved throughout most of the audition circuit. The primary auditory cortex receives direct input from the medial geniculate nucleus of the thalamus and thus is thought to identify the fundamental elements of music, such as pitch and loudness.
An evoked response study of congenitally deaf kittens used local field potentials to measure cortical plasticity in the auditory cortex. These kittens were stimulated and measured against a control (an un-stimulated congenitally deaf cat (CDC)) and normal hearing cats. The field potentials measured for artificially stimulated CDC were eventually much stronger than that of a normal hearing cat.[24] This finding accords with a study by Eckart Altenmuller, in which it was observed that students who received musical instruction had greater cortical activation than those who did not.[25]
The auditory cortex has distinct responses to sounds in the gamma band. When subjects are exposed to three or four cycles of a 40 hertz click, an abnormal spike appears in the EEG data, which is not present for other stimuli. The spike in neuronal activity correlating to this frequency is not restrained to the tonotopic organization of the auditory cortex. It has been theorized that gamma frequencies are resonant frequencies of certain areas of the brain and appear to affect the visual cortex as well.[26] Gamma band activation (25 to 100Hz) has been shown to be present during the perception of sensory events and the process of recognition. In a 2000 study by Kneif and colleagues, subjects were presented with eight musical notes to well-known tunes, such as Yankee Doodle and Frère Jacques. Randomly, the sixth and seventh notes were omitted and an electroencephalogram, as well as a magnetoencephalogram were each employed to measure the neural results. Specifically, the presence of gamma waves, induced by the auditory task at hand, were measured from the temples of the subjects. The omitted stimulus response (OSR)[27] was located in a slightly different position; 7mm more anterior, 13mm more medial and 13mm more superior in respect to the complete sets. The OSR recordings were also characteristically lower in gamma waves as compared to the complete musical set. The evoked responses during the sixth and seventh omitted notes are assumed to be imagined, and were characteristically different, especially in the right hemisphere.[citation needed] The right auditory cortex has long been shown to be more sensitive to tonality (high spectral resolution), while the left auditory cortex has been shown to be more sensitive to minute sequential differences (rapid temporal changes) in sound, such as in speech.[28]
Tonality is represented in more places than just the auditory cortex; one other specific area is the rostromedial prefrontal cortex (RMPFC).[29] A study explored the areas of the brain which were active during tonality processing, using fMRI. The results of this experiment showed preferential blood-oxygen-level-dependent activation of specific voxels in RMPFC for specific tonal arrangements. Though these collections of voxels do not represent the same tonal arrangements between subjects or within subjects over multiple trials, it is interesting and informative that RMPFC, an area not usually associated with audition, seems to code for immediate tonal arrangements in this respect. RMPFC is a subsection of the medial prefrontal cortex, which projects to many diverse areas including the amygdala, and is thought to aid in the inhibition of negative emotion.[30]
Another study has suggested that people who experience 'chills' while listening to music have a higher volume of fibres connecting their auditory cortex to areas associated with emotional processing.[31]
In a study involving dichotic listening to speech, in which one message is presented to the right ear and another to the left, it was found that the participants chose letters with stops (e.g. 'p', 't', 'k', 'b') far more often when presented to the right ear than the left. However, when presented with phonemic sounds of longer duration, such as vowels, the participants did not favor any particular ear.[32] Due to the contralateral nature of the auditory system, the right ear is connected to Wernicke's area, located within the posterior section of the superior temporal gyrus in the left cerebral hemisphere.
Sounds entering the auditory cortex are treated differently depending on whether or not they register as speech. When people listen to speech, according to the strong and weak speech mode hypotheses, they, respectively, engage perceptual mechanisms unique to speech or engage their knowledge of language as a whole.
A Brodmann area is a region of the cerebral cortex, in the human or other primate brain, defined by its cytoarchitecture, or histological structure and organization of cells. The concept was first introduced by the German anatomist Korbinian Brodmann in the early 20th century. Brodmann mapped the human brain based on the varied cellular structure across the cortex and identified 52 distinct regions, which he numbered 1 to 52. These regions, or Brodmann areas, correspond with diverse functions including sensation, motor control, and cognition.
The temporal lobe is one of the four major lobes of the cerebral cortex in the brain of mammals. The temporal lobe is located beneath the lateral fissure on both cerebral hemispheres of the mammalian brain.
The auditory system is the sensory system for the sense of hearing. It includes both the sensory organs and the auditory parts of the sensory system.
The transverse temporal gyri, also called Heschl's gyri or Heschl's convolutions, are gyri found in the area of primary auditory cortex buried within the lateral sulcus of the human brain, occupying Brodmann areas 41 and 42. Transverse temporal gyri are superior to and separated from the planum temporale by Heschl's sulcus. Transverse temporal gyri are found in varying numbers in both the right and left hemispheres of the brain and one study found that this number is not related to the hemisphere or dominance of hemisphere studied in subjects. Transverse temporal gyri can be viewed in the sagittal plane as either an omega shape or a heart shape.
The superior temporal gyrus (STG) is one of three gyri in the temporal lobe of the human brain, which is located laterally to the head, situated somewhat above the external ear.
The planum temporale is the cortical area just posterior to the auditory cortex within the Sylvian fissure. It is a triangular region which forms the heart of Wernicke's area, one of the most important functional areas for language. Original studies on this area found that the planum temporale was one of the most asymmetric regions in the brain, with this area being up to ten times larger in the left cerebral hemisphere than the right.
In physiology, tonotopy is the spatial arrangement of where sounds of different frequency are processed in the brain. Tones close to each other in terms of frequency are represented in topologically neighbouring regions in the brain. Tonotopic maps are a particular case of topographic organization, similar to retinotopy in the visual system.
Multisensory integration, also known as multimodal integration, is the study of how information from the different sensory modalities may be integrated by the nervous system. A coherent representation of objects combining modalities enables animals to have meaningful perceptual experiences. Indeed, multisensory integration is central to adaptive behavior because it allows animals to perceive a world of coherent perceptual entities. Multisensory integration also deals with how different sensory modalities interact with one another and alter each other's processing.
In psycholinguistics, language processing refers to the way humans use words to communicate ideas and feelings, and how such communications are processed and understood. Language processing is considered to be a uniquely human ability that is not produced with the same grammatical understanding or systematicity in even human's closest primate relatives.
A topographic map is the ordered projection of a sensory surface, like the retina or the skin, or an effector system, like the musculature, to one or more structures of the central nervous system. Topographic maps can be found in all sensory systems and in many motor systems.
Cortical deafness is a rare form of sensorineural hearing loss caused by damage to the primary auditory cortex. Cortical deafness is an auditory disorder where the patient is unable to hear sounds but has no apparent damage to the structures of the ear. It has been argued to be as the combination of auditory verbal agnosia and auditory agnosia. Patients with cortical deafness cannot hear any sounds, that is, they are not aware of sounds including non-speech, voices, and speech sounds. Although patients appear and feel completely deaf, they can still exhibit some reflex responses such as turning their head towards a loud sound.
In human neuroanatomy, brain asymmetry can refer to at least two quite distinct findings:
Auditory agnosia is a form of agnosia that manifests itself primarily in the inability to recognize or differentiate between sounds. It is not a defect of the ear or "hearing", but rather a neurological inability of the brain to process sound meaning. While auditory agnosia impairs the understanding of sounds, other abilities such as reading, writing, and speaking are not hindered. It is caused by bilateral damage to the anterior superior temporal gyrus, which is part of the auditory pathway responsible for sound recognition, the auditory "what" pathway.
In neuroscience, the N100 or N1 is a large, negative-going evoked potential measured by electroencephalography ; it peaks in adults between 80 and 120 milliseconds after the onset of a stimulus, and is distributed mostly over the fronto-central region of the scalp. It is elicited by any unpredictable stimulus in the absence of task demands. It is often referred to with the following P200 evoked potential as the "N100-P200" or "N1-P2" complex. While most research focuses on auditory stimuli, the N100 also occurs for visual, olfactory, heat, pain, balance, respiration blocking, and somatosensory stimuli.
In the human brain, the superior temporal sulcus (STS) is the sulcus separating the superior temporal gyrus from the middle temporal gyrus in the temporal lobe of the brain. A sulcus is a deep groove that curves into the largest part of the brain, the cerebrum, and a gyrus is a ridge that curves outward of the cerebrum.
The neuroscience of music is the scientific study of brain-based mechanisms involved in the cognitive processes underlying music. These behaviours include music listening, performing, composing, reading, writing, and ancillary activities. It also is increasingly concerned with the brain basis for musical aesthetics and musical emotion. Scientists working in this field may have training in cognitive neuroscience, neurology, neuroanatomy, psychology, music theory, computer science, and other relevant fields.
The neural encoding of sound is the representation of auditory sensation and perception in the nervous system. The complexities of contemporary neuroscience are continually redefined. Thus what is known of the auditory system has been continually changing. The encoding of sounds includes the transduction of sound waves into electrical impulses along auditory nerve fibers, and further processing in the brain.
Cross modal plasticity is the adaptive reorganization of neurons to integrate the function of two or more sensory systems. Cross modal plasticity is a type of neuroplasticity and often occurs after sensory deprivation due to disease or brain damage. The reorganization of the neural network is greatest following long-term sensory deprivation, such as congenital blindness or pre-lingual deafness. In these instances, cross modal plasticity can strengthen other sensory systems to compensate for the lack of vision or hearing. This strengthening is due to new connections that are formed to brain cortices that no longer receive sensory input.
Temporal envelope (ENV) and temporal fine structure (TFS) are changes in the amplitude and frequency of sound perceived by humans over time. These temporal changes are responsible for several aspects of auditory perception, including loudness, pitch and timbre perception and spatial hearing.
Auditosensory cortex is the part of the auditory system that is associated with the sense of hearing in humans. It occupies the bilateral primary auditory cortex in the temporal lobe of the mammalian brain. The term is used to describe Brodmann area 42 together with the transverse temporal gyri of Heschl. The auditosensory cortex takes part in the reception and processing of auditory nerve impulses, which passes sound information from the thalamus to the brain. Abnormalities in this region are responsible for many disorders in auditory abilities, such as congenital deafness, true cortical deafness, primary progressive aphasia and auditory hallucination.
References
↑ Cf. Pickles, James O. (2012). An Introduction to the Physiology of Hearing (4th ed.). Bingley, UK: Emerald Group Publishing Limited, p. 238.
↑ Kulynych, J. J.; Vladar, K.; Jones, D. W.; Weinberger, D. R. (March 1994). "Gender differences in the normal lateralization of the supratemporal cortex: MRI surface-rendering morphometry of Heschl's gyrus and the planum temporale". Cerebral Cortex. 4 (2): 107–118. doi:10.1093/cercor/4.2.107. ISSN1047-3211. PMID8038562.
↑ Deutsch, Diana (February 2010). "Hearing Music in Ensembles". Physics Today. Vol.63, no.2. p.40. doi:10.1063/1.3326988.
↑ Cant, NB; Benson, CG (June 15, 2003). "Parallel auditory pathways: projection patterns of the different neuronal populations in the dorsal and ventral cochlear nuclei". Brain Res Bull. 60 (5–6): 457–74. doi:10.1016/S0361-9230(03)00050-9. PMID12787867. S2CID42563918.
↑ Dinh, L; Nguyen T; Salgado H; Atzori M (2009). "Norepinephrine homogeneously inhibits alpha-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate- (AMPAR-) mediated currents in all layers of the temporal cortex of the rat". Neurochem Res. 34 (11): 1896–906. doi:10.1007/s11064-009-9966-z. PMID19357950. S2CID25255160.
↑ Lauter, Judith L; P Herscovitch; C Formby; ME Raichle (1985). "Tonotopic organization in human auditory cortex revealed by positron emission tomography". Hearing Research. 20 (3): 199–205. doi:10.1016/0378-5955(85)90024-3. PMID3878839. S2CID45928728.
↑ Klinke, Rainer; Kral, Andrej; Heid, Silvia; Tillein, Jochen; Hartmann, Rainer (September 10, 1999). "Recruitment of the auditory cortex in congenitally deaf cats by long-term cochlear electrostimulation". Science. 285 (5434): 1729–33. doi:10.1126/science.285.5434.1729. PMID10481008. S2CID38985173.
↑ Tallon-Baudry, C.; Bertrand, O. (April 1999). "Oscillatory gamma activity in humans and its role in object representation". Trends in Cognitive Sciences. 3 (4): 151–162. doi:10.1016/S1364-6613(99)01299-1. PMID10322469. S2CID1308261.
↑ Busse, L; Woldorff, M (April 2003). "The ERP omitted stimulus response to "no-stim" events and its implications for fast-rate event-related fMRI designs". NeuroImage. 18 (4): 856–864. doi:10.1016/s1053-8119(03)00012-0. PMID12725762. S2CID25351923.
↑ Cassel, M. D.; Wright, D. J. (September 1986). "Topography of projections from the medial prefrontal cortex to the amygdala in the rat". Brain Research Bulletin. 17 (3): 321–333. doi:10.1016/0361-9230(86)90237-6. PMID2429740. S2CID22826730.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.