| Temporal lobe | |
|---|---|
Lobes of the human brain (temporal lobe is shown in green) | |
 Section of brain showing upper surface of temporal lobe. | |
| Details | |
| Part of | Cerebrum |
| Artery | |
| Vein | |
| Identifiers | |
| Latin | lobus temporalis |
| MeSH | D013702 |
| NeuroNames | 125 |
| NeuroLex ID | birnlex_1160 |
| TA98 | A14.1.09.136 |
| TA2 | 5488 |
| FMA | 61825 |
| Anatomical terms of neuroanatomy | |
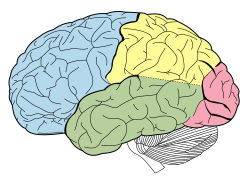
The temporal lobe is one of the four major lobes of the cerebral cortex in the brain of mammals. The temporal lobe is located beneath the lateral fissure on both cerebral hemispheres of the mammalian brain. [3]
Contents
- Structure
- Function
- Visual memories
- Processing sensory input
- Language recognition
- New memories
- Clinical significance
- Unilateral temporal lesion
- Dominant hemisphere
- Non-dominant hemisphere
- Bitemporal lesions (additional features)
- Damage
- Disorders
- See also
- References
- External links
The temporal lobe is involved in processing sensory input into derived meanings for the appropriate retention of visual memory, language comprehension, and emotion association. [4] : 21 Temporal refers to the head's temples.
