The cerebellum develops in a rostro-caudal manner, with rostral regions in the midline giving rise to the vermis, and caudal regions developing into the cerebellar hemispheres.[2] By 4 months of prenatal development, the vermis becomes fully foliated, while development of the hemispheres lags by 30–60 days.[3] Postnatally, proliferation and organization of the cellular components of the cerebellum continues, with completion of the foliation pattern by 7 months of life[4] and final migration, proliferation, and arborization of cerebellar neurons by 20 months.[5]
The vermis is intimately associated with all regions of the cerebellar cortex, which can be divided into three functional parts, each having distinct connections with the brain and spinal cord. These regions are the vestibulocerebellum, which is responsible primarily for the control of eye movements; the spinocerebellum, involved in fine tune body and limb movement; and the cerebrocerebellum, which is associated with planning, initiation and timing of movements.[8]
Structure
Anterior surface of cerebellum. The vermis is highlighted in red.
The vermis is the unpaired, median portion of the cerebellum that connects the two hemispheres.[9] Both the vermis and the hemispheres are composed of lobules formed by groups of folia. There are nine lobules of the vermis: lingula, central lobule, culmen, clivus, folium of the vermis, tuber, pyramid, uvula and nodule.[9] These lobules are often difficult to observe during human anatomy classes and may vary in size, shape and number of folia. It has been shown that folia of the cerebellum exhibit frequent variations in form, number and arrangement between individuals.[9]
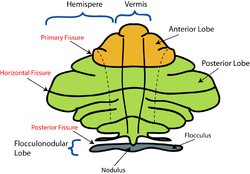
Lobe anatomy
Schematic representation of the major anatomical subdivisions of the cerebellum.
The lingula is the first lobule of the upper portion of the vermis on the superoinferior axis and pertains to the paleocerebellum together with the central lobule, culmen, pyramid and uvula. It is separated from the central lobule by the pre-central fissure. The central lobule is the second lobule of the upper portion of the vermis on the superoinferior axis. The culmen is the third and largest lobule of the upper portion of the vermis on the superoinferior axis. It is separated from the declive by the primary fissure and is related with the anterior quadrangular lobule of the hemisphere. The pyramid is the seventh lobule of the vermis on the superoinferior axis. It is separated from the tuber and uvula by the pre-pyramidal and secondary fissures, respectively.[9] This lobule is related with the biventral lobule of the hemisphere. The uvula is the second largest lobule, following the culmen. It pertains to the paleocerebellum and is separated from the nodule by the posterolateral fissure.[9]
Spinocerebellum
The spinocerebellum receives proprioception input from the dorsal columns of the spinal cord (including the spinocerebellar tract) and from the trigeminal nerve, as well as from visual and auditory systems. It sends fibers to deep cerebellar nuclei that, in turn, project to both the cerebral cortex and the brain stem, thus providing modulation of descending motor systems.[8] This region comprises the vermis and intermediate parts of the cerebellar hemispheres. Sensory information from the periphery and from the primary motor and somatosensory cortex terminate in this region. Purkinje cells of the vermis project to the fastigial nucleus, controlling the axial and proximal musculature involved in the execution of limb movements.[10]Purkinje cells in the intermediate zone of the spinocerebellum project to the interposed nuclei, which control the distal musculature components of the descending motor pathways needed for limb movement. Both of these nuclei include projections to the motor cortex in the cerebrum.[10]
Joubert syndrome (JS) is one of the most commonly diagnosed syndromes associated with the molar tooth sign (MTS),[13] or hypoplasia/dysplasia of the cerebellar vermis accompanied by brainstem abnormalities. JS is defined clinically by features of hypotonia in infancy with later development of ataxia, developmental delays, mental retardation, abnormal breathing patterns, abnormal eye movements specific to oculomotor apraxia, or the presence of the MTS on the cranial MRI.[14][15]JS is an autosomal recessive condition with an estimated prevalence of 1: 100,000.[16]
Dandy Walker malformation
Dandy Walker malformation is a relatively common congenital brain malformation with a prevalence of 1:30,000 live births.[17] Dandy Walker malformation is characterized by enlarged posterior fossa and in which the cerebellar vermis is completely absent, or present in a rudimentary form, sometimes rotated accompanied by an elevation of the fourth ventricle. It is also commonly associated with dysplasias of brainstem nuclei.[18] DWM has been reported to be in association with a wide array of chromosomal anomalies, including trisomy 18, trisomy 9, and trisomy 13. Surveys suggest that prenatal exposure to teratogens such as rubella or alcohol are correlated with development of Dandy Walker malformation.[19][20]
Hypoplasia and other structural alterations of the vermis have been identified in many patients with autism spectrum disorder (ASD). While the exact nature and extent of the impacts ASD has on the vermis remain in question, it has also been shown that other injuries and malformations of the vermis sometimes produce symptoms closely analogous to ASD. Furthermore, several genetic syndromes known to cause autism (such as fragile X syndrome) have also been shown to cause damage to the vermis.[24]
Damage
Lesions to the vermis commonly give rise to clinical depression, inappropriate emotional displays (e.g. unwarranted giggling) in addition to movement disorders. [citation needed]
Comparative anatomy
Early neurophysiologists suggest that retinal and inertial signals were selected for about 450 million years ago by primitive brainstem-cerebellar circuitry because of their relationship with the environment.[25] Microscopically, it is evident that Purkinje cell precursors arose from granule cells, first forming in irregular patterns, then progressively becoming organized in a layered fashion. Evolutionarily, the Purkinje cells then developed extensive dendritic trees that increasingly became confined to a single plane, through which the axons of granule cells threaded, eventually forming a neuronal grid of right angles.[25] The origin of the cerebellum is in close association with that of the nuclei of the vestibular cranial nerve and lateral line nerves, perhaps suggesting that this part of the cerebellum originated as a means of carrying out transformations of the coordinate system from input data of the vestibular organ and the lateral line organs.[26] This suggests that the function of the cerebellum evolved as a mode of computing and representing an image relating to the position of the body in space. The cerebellar vermis evolved in conjunction with the hemispheres; this is seen in lampreys and higher vertebrates.[27]
In fish
In vertebrates, the cerebellar vermis develops between two bilaterally symmetrical formations located dorsal to the upper end of the medulla oblongata, or rhombencephalon. This is the region of termination for the fibers of the vestibular nerve and lateral line nerves; thus, these are the oldest afferent paths to the cerebellum and cerebellar vermis.[27] In bony fish, or teleosts, it has been proposed that the cerebellar auricles, which receive a large amount of input from the vestibulolateral line system, constitute the vestibulocerebellum and are homologues of the flocculonodular lobe of higher vertebrates along with the corpus cerebelli, which receives spinocerebellar and tectocerebellar fibers. The labyrinth and the lateral line organs of lampreys have structural and functional similarity. An important difference between the two structures is that the arrangement of the lateral line organs are such that they are sensitive to relative motion of the fluid surrounding the animal, whereas the labyrinths, having very similar sensing mechanisms, are sensitive to endolymph, providing information concerning the animal's own equilibrium of the body and orientation in space.[27]
↑ Cho, K. H.; Rodriguez-Vazquez, J. F.; Kim, J. H.; Abe, H.; Murakami, G.; Cho, B. H. (2011). "Early fetal development of the human cerebellum". Surgical and Radiologic Anatomy. 33 (6): 523–530. doi:10.1007/s00276-011-0796-8. PMID21380713. S2CID25451924.
↑ Parisia, M.; Dobynsb, W. (2003). "Human malformations of the midbrain and hindbrain: review and proposed classification scheme". Molecular Genetics and Metabolism. 80 (1–2): 36–53. doi:10.1016/j.ymgme.2003.08.010. PMID14567956.
↑ Robinson AJ, Blaser S, Toi A, etal. (2007). "The fetal cerebellar vermis: assessment for abnormal development by ultrasonography and magnetic resonance imaging". Ultrasound Quarterly. 23 (3): 211–223. doi:10.1097/ruq.0b013e31814b162c. PMID17805192. S2CID23068656.
1 2 3 Ghez C, Fahn S (1985). "The cerebellum". In Kandel ER, Schwartz JH (eds.). Principles of Neural Science (2nded.). New York: Elsevier. pp.502–522.
↑ J.M. Saraiva; M. Baraitser (1992). "Joubert syndrome: a review". American Journal of Medical Genetics. 43 (4): 726–731. doi:10.1002/ajmg.1320430415. PMID1341417.
↑ B.L. Maria; E. Boltshauser; S.C. Palmer; T.X. Tran (1999). "Clinical features and revised diagnostic criteria in Joubert syndrome". Child Neurology. 14 (9): 583–590. doi:10.1177/088307389901400906. PMID10488903. S2CID7410607.
↑ D.B. Flannery; J.G. Hudson (1994). A survey of Joubert syndrome. David W. Smith Workshop.
↑ Osenbach, R.K.; Menezes, A.H. (1992). "Diagnosis and management of the Dandy-Walker malformation: 30 years of experience". Pediatric Neurosurgery. 18 (4): 179–89. doi:10.1159/000120660. PMID1472430.
↑ Kapur, R.; Mahony, B.; Finch, L.; Siebert J. (2009). "Normal and Abnormal Anatomy of the Cerebellar Vermis in Midgestational Human Fetuses". Birth Defects Research. 85 (8): 700–709. doi:10.1002/bdra.20589. PMID19441098.
↑ S.K. Clarren; J. Alvord; S.M. Sumi (1978). "Brain malformations related to prenatal exposure to ethanol". Journal of Pediatrics. 92 (1): 64–67. doi:10.1016/S0022-3476(78)80072-9. PMID619080.
↑ S.P. Toelle; C. Yalcinkaya; N. Kocer; T. Deonna; W.C.G. Overweg-Plandsoen; T. Bast; R. Kalmanchey; P. Barsi; J.F.L. Schneider; A. Capone Mori; E. Boltshauser (2002). "Rhombencephalosynapsis: clinical findings and neuroimaging in 9 children". Neuropediatrics. 33 (4): 209–214. doi:10.1055/s-2002-34498. PMID12368992. S2CID32510022.
1 2 Nieuwenhuys, R.; Voogd, J.; van Huijzen, C. (1988). The Human Central Nervous System: A Synopsis and Atlas (3rded.). Heidelberg: Springer-Verlag.
↑ Butler, A.B.; Hodos, W. (1996). "12: The Cerebellum". Comparative Vertebrate Neuroanatomy: Evolution and Adaptation. New York: Wiley-Liss. pp.180–197.
1 2 3 Ariens, K.; C.U.; Huber, G.C.; Crosby, E.C. (1960). The Comparative Anatomy of the Nervous System of Vertebrates, Including Man. Vol.3. New York: Hafner.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.