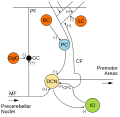
| Basket cell | |
|---|---|
 Transverse section of a cerebellar folium (Basket cell labeled at bottom left) | |
| Details | |
| Location | Cerebellum |
| Shape | multipolar |
| Function | Inhibitory interneuron |
| Neurotransmitter | GABA |
| Presynaptic connections | Parallel fibers |
| Postsynaptic connections | Purkinje cells |
| Identifiers | |
| NeuroLex ID | nifext_160 |
| Anatomical terms of neuroanatomy | |
Basket cells are inhibitory GABAergic interneurons of the brain, found throughout different regions of the cortex and cerebellum. [1]