Related Research Articles

The visual cortex of the brain is the area of the cerebral cortex that processes visual information. It is located in the occipital lobe. Sensory input originating from the eyes travels through the lateral geniculate nucleus in the thalamus and then reaches the visual cortex. The area of the visual cortex that receives the sensory input from the lateral geniculate nucleus is the primary visual cortex, also known as visual area 1 (V1), Brodmann area 17, or the striate cortex. The extrastriate areas consist of visual areas 2, 3, 4, and 5.
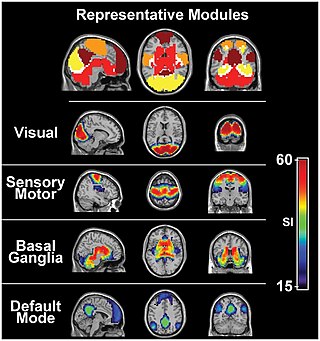
The sensory nervous system is a part of the nervous system responsible for processing sensory information. A sensory system consists of sensory neurons, neural pathways, and parts of the brain involved in sensory perception and interoception. Commonly recognized sensory systems are those for vision, hearing, touch, taste, smell, balance and visceral sensation. Sense organs are transducers that convert data from the outer physical world to the realm of the mind where people interpret the information, creating their perception of the world around them.
Stimulus modality, also called sensory modality, is one aspect of a stimulus or what is perceived after a stimulus. For example, the temperature modality is registered after heat or cold stimulate a receptor. Some sensory modalities include: light, sound, temperature, taste, pressure, and smell. The type and location of the sensory receptor activated by the stimulus plays the primary role in coding the sensation. All sensory modalities work together to heighten stimuli sensation when necessary.

David Hunter Hubel was an American Canadian neurophysiologist noted for his studies of the structure and function of the visual cortex. He was co-recipient with Torsten Wiesel of the 1981 Nobel Prize in Physiology or Medicine, for their discoveries concerning information processing in the visual system. For much of his career, Hubel worked as the Professor of Neurobiology at Johns Hopkins University and Harvard Medical School. In 1978, Hubel and Wiesel were awarded the Louisa Gross Horwitz Prize from Columbia University. In 1983, Hubel received the Golden Plate Award of the American Academy of Achievement.
The receptive field, or sensory space, is a delimited medium where some physiological stimuli can evoke a sensory neuronal response in specific organisms.
Visual processing is a term that refers to the brain's ability to use and interpret visual information from the world. The process of converting light energy into a meaningful image is a complex process that is facilitated by numerous brain structures and higher level cognitive processes.

The inferior temporal gyrus is one of three gyri of the temporal lobe and is located below the middle temporal gyrus, connected behind with the inferior occipital gyrus; it also extends around the infero-lateral border on to the inferior surface of the temporal lobe, where it is limited by the inferior sulcus. This region is one of the higher levels of the ventral stream of visual processing, associated with the representation of objects, places, faces, and colors. It may also be involved in face perception, and in the recognition of numbers and words.

In neurobiology, lateral inhibition is the capacity of an excited neuron to reduce the activity of its neighbors. Lateral inhibition disables the spreading of action potentials from excited neurons to neighboring neurons in the lateral direction. This creates a contrast in stimulation that allows increased sensory perception. It is also referred to as lateral antagonism and occurs primarily in visual processes, but also in tactile, auditory, and even olfactory processing. Cells that utilize lateral inhibition appear primarily in the cerebral cortex and thalamus and make up lateral inhibitory networks (LINs). Artificial lateral inhibition has been incorporated into artificial sensory systems, such as vision chips, hearing systems, and optical mice. An often under-appreciated point is that although lateral inhibition is visualised in a spatial sense, it is also thought to exist in what is known as "lateral inhibition across abstract dimensions." This refers to lateral inhibition between neurons that are not adjacent in a spatial sense, but in terms of modality of stimulus. This phenomenon is thought to aid in colour discrimination.
Neural coding is a neuroscience field concerned with characterising the hypothetical relationship between the stimulus and the neuronal responses, and the relationship among the electrical activities of the neurons in the ensemble. Based on the theory that sensory and other information is represented in the brain by networks of neurons, it is believed that neurons can encode both digital and analog information.
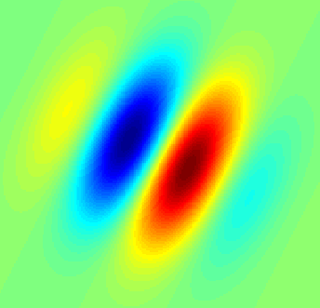
A simple cell in the primary visual cortex is a cell that responds primarily to oriented edges and gratings. These cells were discovered by Torsten Wiesel and David Hubel in the late 1950s.
In neuroanatomy, topographic map is the ordered projection of a sensory surface or an effector system to one or more structures of the central nervous system. Topographic maps can be found in all sensory systems and in many motor systems.
Complex cells can be found in the primary visual cortex (V1), the secondary visual cortex (V2), and Brodmann area 19 (V3).

The colour centre is a region in the brain primarily responsible for visual perception and cortical processing of colour signals received by the eye, which ultimately results in colour vision. The colour centre in humans is thought to be located in the ventral occipital lobe as part of the visual system, in addition to other areas responsible for recognizing and processing specific visual stimuli, such as faces, words, and objects. Many functional magnetic resonance imaging (fMRI) studies in both humans and macaque monkeys have shown colour stimuli to activate multiple areas in the brain, including the fusiform gyrus and the lingual gyrus. These areas, as well as others identified as having a role in colour vision processing, are collectively labelled visual area 4 (V4). The exact mechanisms, location, and function of V4 are still being investigated.

The fusiform face area is a part of the human visual system that is specialized for facial recognition. It is located in the inferior temporal cortex (IT), in the fusiform gyrus.
A hypercomplex cell is a type of visual processing neuron in the mammalian cerebral cortex. Initially discovered by David Hubel and Torsten Wiesel in 1965, hypercomplex cells are defined by the property of end-stopping, which is a decrease in firing strength with increasingly larger stimuli. The sensitivity to stimulus length is accompanied by selectivity for the specific orientation, motion, and direction of stimuli. For example, a hypercomplex cell may only respond to a line at 45˚ that travels upward. Elongating the line would result in a proportionately weaker response. Ultimately, hypercomplex cells can provide a means for the brain to visually perceive corners and curves in the environment by identifying the ends of a given stimulus.
A sensory map is an area of the brain which responds to sensory stimulation, and are spatially organized according to some feature of the sensory stimulation. In some cases the sensory map is simply a topographic representation of a sensory surface such as the skin, cochlea, or retina. In other cases it represents other stimulus properties resulting from neuronal computation and is generally ordered in a manner that reflects the periphery. An example is the somatosensory map which is a projection of the skin's surface in the brain that arranges the processing of tactile sensation. This type of somatotopic map is the most common, possibly because it allows for physically neighboring areas of the brain to react to physically similar stimuli in the periphery or because it allows for greater motor control.
An organism is said to be sexually dimorphic when male and female conspecifics have anatomical differences in features such as body size, coloration, or ornamentation, but disregarding differences of reproductive organs. Sexual dimorphism is usually a product of sexual selection, with female choice leading to elaborate male ornamentation and male-male competition leading to the development of competitive weaponry. However, evolutionary selection also acts on the sensory systems that receivers use to perceive external stimuli. If the benefits of perception to one sex or the other are different, sex differences in sensory systems can arise. For example, female production of signals used to attract mates can put selective pressure on males to improve their ability to detect those signals. As a result, only males of this species will evolve specialized mechanisms to aid in detection of the female signal. This article uses examples of sex differences in the olfactory, visual, and auditory systems of various organisms to show how sex differences in sensory systems arise when it benefits one sex and not the other to have enhanced perception of certain external stimuli. In each case, the form of the sex difference reflects the function it serves in terms of enhanced reproductive success.
Feature detection is a process by which the nervous system sorts or filters complex natural stimuli in order to extract behaviorally relevant cues that have a high probability of being associated with important objects or organisms in their environment, as opposed to irrelevant background or noise.

Orientation columns are organized regions of neurons that are excited by visual line stimuli of varying angles. These columns are located in the primary visual cortex (V1) and span multiple cortical layers. The geometry of the orientation columns are arranged in slabs that are perpendicular to the surface of the primary visual cortex.
Binocular neurons are neurons in the visual system that assist in the creation of stereopsis from binocular disparity. They have been found in the primary visual cortex where the initial stage of binocular convergence begins. Binocular neurons receive inputs from both the right and left eyes and integrate the signals together to create a perception of depth.
References
- ↑ Sakai, Kuniyoshi; Miyashita, Yasushi. Neuronal tuning to learned complex forms in vision. NeuroReport 1994, 5:829-832.
- 1 2 Matteo Carandini, Jonathan B. Demb, Valerio Mante, David J. Tolhurst, Yang Dan, Bruno A. Olshausen, Jack L. Gallant and Nicole C. Rust. Do we know what the early visual system does? Journal of Neuroscience 25:10577-10597.
- ↑ Grill-Spector, Kalanit; Witthoft, Nathan. Deos the Bairn Not Raed Ervey Lteter by Istlef, but the Wrod as a Wlohe? Neuron 2009, 62:161-162.
- ↑ Teller. Linking propositions. Vision Research 1984,24:1233-2346.
- 1 2 3 Hubel, D. H.; Wiesel, T. N. Receptive Fields of Single Neurones in the Cat's Striate Cortex. J. Physiol. 1959, 148:574-591.
- ↑ Wurtz, Robert H. Recounting the impact of Hubel and Wiesel. J. Physiol. 2009, 587:2817-2823.
- ↑ www.nobelprize.org
- 1 2 Riesenhuber, Maximilian; Poggio, Tomaso. Neural mechanisms of object recognition. Current Opinion in Neurobiology 2002, 12:162-168.
- 1 2 3 4 5 6 7 8 9 Kolb, B.; Whishaw, I. Q. Fundamentals of Human Neuropsychology (2003). New York, NY: Worth Publishers.
- 1 2 Sakai, K.; Naya, Y.; Miyashita, Y. Neuronal tuning and associative mechanisms in form representation. Learning and Memory 1994, 1:83-105.
- ↑ Adamson & Troiani. Distinct and overlapping fusiform activation to faces and food. Neuroimage 2018, 174: 393-406
- ↑ Gauthier, I.; Skudlarski, P.; Gore, J. C.; Anderson, A. W. Expertise for cars and birds recruits brain areas involved in face recognition. Nat. Neurosci. 2001, 3:191-197.
- 1 2 Gauthier, I.; Tarr, M. J. Unraveling mechanisms for expert object recognition: Bridging brain activity and behavior. Journal of Experimental Psychology: Human Perception and Performance 2002, 28(2):431-446.
- ↑ Curran, I.; Tanaka, J.; Weiskopf, D. An electrophysiological comparison of visual categorization and recognition memory. Cognitive, Affective, & Behavioral Neuroscience 2002, 18(2):1-18.