
A bone is a rigid organ that constitutes part of the skeleton in most vertebrate animals. Bones protect the various other organs of the body, produce red and white blood cells, store minerals, provide structure and support for the body, and enable mobility. Bones come in a variety of shapes and sizes and have complex internal and external structures. They are lightweight yet strong and hard and serve multiple functions.

Cartilage is a resilient and smooth type of connective tissue. Semi-transparent and non-porous, it is usually covered by a tough and fibrous membrane called perichondrium. In tetrapods, it covers and protects the ends of long bones at the joints as articular cartilage, and is a structural component of many body parts including the rib cage, the neck and the bronchial tubes, and the intervertebral discs. In other taxa, such as chondrichthyans and cyclostomes, it constitutes a much greater proportion of the skeleton. It is not as hard and rigid as bone, but it is much stiffer and much less flexible than muscle. The matrix of cartilage is made up of glycosaminoglycans, proteoglycans, collagen fibers and, sometimes, elastin. It usually grows quicker than bone.

Bone healing, or fracture healing, is a proliferative physiological process in which the body facilitates the repair of a bone fracture.

The tibia, also known as the shinbone or shankbone, is the larger, stronger, and anterior (frontal) of the two bones in the leg below the knee in vertebrates ; it connects the knee with the ankle. The tibia is found on the medial side of the leg next to the fibula and closer to the median plane. The tibia is connected to the fibula by the interosseous membrane of leg, forming a type of fibrous joint called a syndesmosis with very little movement. The tibia is named for the flute tibia. It is the second largest bone in the human body, after the femur. The leg bones are the strongest long bones as they support the rest of the body.

An epiphysis is one of the rounded ends or tips of a long bone that ossify from one or more secondary centers of ossification. Between the epiphysis and diaphysis lies the metaphysis, including the epiphyseal plate. During formation of the secondary ossification center, vascular canals stemming from the perichondrium invade the epiphysis, supplying nutrients to the developing secondary centers of ossification. At the joint, the epiphysis is covered with articular cartilage; below that covering is a zone similar to the epiphyseal plate, known as subchondral bone. The epiphysis is mostly found in mammals but it is also present in some lizards. However, the secondary center of ossification may have evolved multiple times, having been found in the Jurassic sphenodont Sapheosaurus as well as in the therapsid Niassodon mfumukasi.

The long bones are those that are longer than they are wide. They are one of five types of bones: long, short, flat, irregular and sesamoid. Long bones, especially the femur and tibia, are subjected to most of the load during daily activities and they are crucial for skeletal mobility. They grow primarily by elongation of the diaphysis, with an epiphysis at each end of the growing bone. The ends of epiphyses are covered with hyaline cartilage. The longitudinal growth of long bones is a result of endochondral ossification at the epiphyseal plate. Bone growth in length is stimulated by the production of growth hormone (GH), a secretion of the anterior lobe of the pituitary gland.
Hyaline cartilage is the glass-like (hyaline) and translucent cartilage found on many joint surfaces. It is also most commonly found in the ribs, nose, larynx, and trachea. Hyaline cartilage is pearl-gray in color, with a firm consistency and has a considerable amount of collagen. It contains no nerves or blood vessels, and its structure is relatively simple.
Chondrocytes are the only cells found in healthy cartilage. They produce and maintain the cartilaginous matrix, which consists mainly of collagen and proteoglycans. Although the word chondroblast is commonly used to describe an immature chondrocyte, the term is imprecise, since the progenitor of chondrocytes can differentiate into various cell types, including osteoblasts.
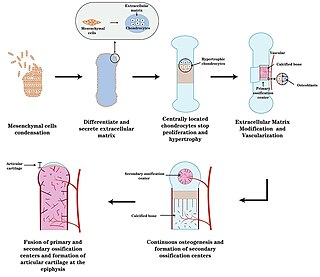
Endochondral ossification is one of the two essential pathways by which bone tissue is produced during fetal development of the mammalian skeletal system, the other pathway being intramembranous ossification. Both endochondral and intramembranous processes initiate from a precursor mesenchymal tissue, but their transformations into bone are different. In intramembranous ossification, mesenchymal tissue is directly converted into bone. On the other hand, endochondral ossification starts with mesenchymal tissue turning into an intermediate cartilage stage, which is eventually substituted by bone.

In osteology, the osteon or haversian system is the fundamental functional unit of much compact bone. Osteons are roughly cylindrical structures that are typically between 0.25 mm and 0.35 mm in diameter. Their length is often hard to define, but estimates vary from several millimeters to around 1 centimeter. They are present in many bones of most mammals and some bird, reptile, and amphibian species.
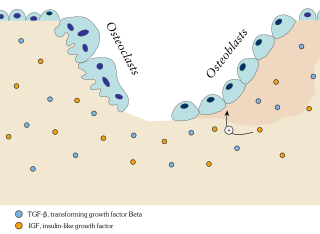
Ossification in bone remodeling is the process of laying down new bone material by cells named osteoblasts. It is synonymous with bone tissue formation. There are two processes resulting in the formation of normal, healthy bone tissue: Intramembranous ossification is the direct laying down of bone into the primitive connective tissue (mesenchyme), while endochondral ossification involves cartilage as a precursor.

Chondroblasts, or perichondrial cells, is the name given to mesenchymal progenitor cells in situ which, from endochondral ossification, will form chondrocytes in the growing cartilage matrix. Another name for them is subchondral cortico-spongious progenitors. They have euchromatic nuclei and stain by basic dyes.

Chondrogenesis is the biological process through which cartilage tissue is formed and developed. This intricate and tightly regulated cellular differentiation pathway plays a crucial role in skeletal development, as cartilage serves as a fundamental component of the embryonic skeleton. The term "chondrogenesis" is derived from the Greek words "chondros," meaning cartilage, and "genesis," meaning origin or formation.

The epiphyseal plate, epiphysial plate, physis, or growth plate is a hyaline cartilage plate in the metaphysis at each end of a long bone. It is the part of a long bone where new bone growth takes place; that is, the whole bone is alive, with maintenance remodeling throughout its existing bone tissue, but the growth plate is the place where the long bone grows longer.

Short bones are designated as those bones that are more or less equal in length, width, and thickness. They include the tarsals in the ankle and the carpals in the wrist. They are one of five types of bones: short, long, flat, irregular and sesamoid. Most short bones are named according to their shape as they exhibit a variety of complex morphological features
In histology, a lacuna is a small space, containing an osteocyte in bone, or chondrocyte in cartilage.

Chondroblastoma is a rare, benign, locally aggressive bone tumor that typically affects the epiphyses or apophyses of long bones. It is thought to arise from an outgrowth of immature cartilage cells (chondroblasts) from secondary ossification centers, originating from the epiphyseal plate or some remnant of it.

Keutel syndrome (KS) is a rare autosomal recessive genetic disorder characterized by abnormal diffuse cartilage calcification, hypoplasia of the mid-face, peripheral pulmonary stenosis, hearing loss, short distal phalanges (tips) of the fingers and mild mental retardation. Individuals with KS often present with peripheral pulmonary stenosis, brachytelephalangism, sloping forehead, midface hypoplasia, and receding chin. It is associated with abnormalities in the gene coding for matrix gla protein, MGP. Being an autosomal recessive disorder, it may be inherited from two unaffected, abnormal MGP-carrying parents. Thus, people who inherit two affected MGP alleles will likely inherit KS.
Primary bone is the first bone tissue that appears in embryonic development and in fracture repair. It is characterized by its random position of collagen fibers. In most places in adults this tissue is replaced by secondary bone tissue except, for example, near the sutures of calvara or tooth sockets. The secondary bones have lower amounts of osteocytes so primary bone is much more easily penetrated by x-ray.
Chondrocalcin comes from a family of pro-collagen molecules. This protein is found in the bone and is involved in calcium-binding, which occurs in order to harden the cartilage. Another one of its roles is to assemble the triple collagen helix, which is predominately entails the linkage of glycine and proline amino acids and then the twisting of those linkages. Chondrocalcin is important because cartilage calcification of the growth plate is one of the main occurrences in endochondral bone formation. Because of its importance, it is one of the most highly created polypeptides in human cartilage. This calcium-binding protein comes from chondrocytes, which are cells that produce and maintain cartilage. Some examples of chondrocytes include collagen and proteoglycans. The chondrocytes that produce chondrocalcin are typically found in growing bone matrices that have not yet matured. These immature bone matrices are found in the epiphyseal plate at both the lower hypertrophic zone and the longitudinal septa of the cartilaginous matrix. One study was conducted on fetal cartilage discovered that the chondrocalcin protein exists as a dimer with 35,000 g/mol subunits. The reason why fetal cartilage was used is because chondrocalcin cannot be detected in mature bone. Another investigation was conducted to determine the mechanism behind calcium binding in chondrocalcin. It was discovered that along with cartilage building, the protein also had a role in cartilage destruction.


