Cladistics is an approach to biological classification in which organisms are categorized in groups ("clades") based on hypotheses of most recent common ancestry. The evidence for hypothesized relationships is typically shared derived characteristics (synapomorphies) that are not present in more distant groups and ancestors. However, from an empirical perspective, common ancestors are inferences based on a cladistic hypothesis of relationships of taxa whose character states can be observed. Theoretically, a last common ancestor and all its descendants constitute a (minimal) clade. Importantly, all descendants stay in their overarching ancestral clade. For example, if the terms worms or fishes were used within a strict cladistic framework, these terms would include humans. Many of these terms are normally used paraphyletically, outside of cladistics, e.g. as a 'grade', which are fruitless to precisely delineate, especially when including extinct species. Radiation results in the generation of new subclades by bifurcation, but in practice sexual hybridization may blur very closely related groupings.

A tetrapod is any four-limbed vertebrate animal of the superclass Tetrapoda. Tetrapods include all extant and extinct amphibians and amniotes, with the latter in turn evolving into two major clades, the sauropsids and synapsids. Some tetrapods, such as snakes, legless lizards, and caecilians, have evolved to become limbless via mutations of the Hox gene. Nevertheless, these limbless groups still qualify as tetrapods through their ancestry, and some retain a pair of vestigial spurs that are remnants of the hindlimbs.

Convergent evolution is the independent evolution of similar features in species of different periods or epochs in time. Convergent evolution creates analogous structures that have similar form or function but were not present in the last common ancestor of those groups. The cladistic term for the same phenomenon is homoplasy. The recurrent evolution of flight is a classic example, as flying insects, birds, pterosaurs, and bats have independently evolved the useful capacity of flight. Functionally similar features that have arisen through convergent evolution are analogous, whereas homologous structures or traits have a common origin but can have dissimilar functions. Bird, bat, and pterosaur wings are analogous structures, but their forelimbs are homologous, sharing an ancestral state despite serving different functions.

In biology, homology is similarity due to shared ancestry between a pair of structures or genes in different taxa. A common example of homologous structures is the forelimbs of vertebrates, where the wings of bats and birds, the arms of primates, the front flippers of whales, and the forelegs of four-legged vertebrates like dogs and crocodiles are all derived from the same ancestral tetrapod structure. Evolutionary biology explains homologous structures adapted to different purposes as the result of descent with modification from a common ancestor. The term was first applied to biology in a non-evolutionary context by the anatomist Richard Owen in 1843. Homology was later explained by Charles Darwin's theory of evolution in 1859, but had been observed before this, from Aristotle onwards, and it was explicitly analysed by Pierre Belon in 1555.
Comparative anatomy is the study of similarities and differences in the anatomy of different species. It is closely related to evolutionary biology and phylogeny.
Evolutionary taxonomy, evolutionary systematics or Darwinian classification is a branch of biological classification that seeks to classify organisms using a combination of phylogenetic relationship, progenitor-descendant relationship, and degree of evolutionary change. This type of taxonomy may consider whole taxa rather than single species, so that groups of species can be inferred as giving rise to new groups. The concept found its most well-known form in the modern evolutionary synthesis of the early 1940s.

The triune brain is a model of the evolution of the vertebrate forebrain and behavior, proposed by the American physician and neuroscientist Paul D. MacLean in the 1960s. The triune brain consists of the reptilian complex, the paleomammalian complex, and the neomammalian complex (neocortex), viewed each as independently conscious, and as structures sequentially added to the forebrain in the course of evolution. According to the model, the basal ganglia are in charge of primal instincts, the limbic system is in charge of emotions, and the neocortex is responsible for objective or rational thoughts.

In evolutionary developmental biology, heterochrony is any genetically controlled difference in the timing, rate, or duration of a developmental process in an organism compared to its ancestors or other organisms. This leads to changes in the size, shape, characteristics and even presence of certain organs and features. It is contrasted with heterotopy, a change in spatial positioning of some process in the embryo, which can also create morphological innovation. Heterochrony can be divided into intraspecific heterochrony, variation within a species, and interspecific heterochrony, phylogenetic variation, i.e. variation of a descendant species with respect to an ancestral species.
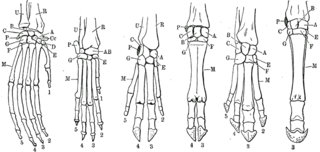
A forelimb or front limb is one of the paired articulated appendages (limbs) attached on the cranial (anterior) end of a terrestrial tetrapod vertebrate's torso. With reference to quadrupeds, the term foreleg or front leg is often used instead. In bipedal animals with an upright posture, the term upper limb is often used.
Evolutionary neuroscience is the scientific study of the evolution of nervous systems. Evolutionary neuroscientists investigate the evolution and natural history of nervous system structure, functions and emergent properties. The field draws on concepts and findings from both neuroscience and evolutionary biology. Historically, most empirical work has been in the area of comparative neuroanatomy, and modern studies often make use of phylogenetic comparative methods. Selective breeding and experimental evolution approaches are also being used more frequently.
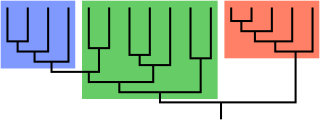
A grade is a taxon united by a level of morphological or physiological complexity. The term was coined by British biologist Julian Huxley, to contrast with clade, a strictly phylogenetic unit.

A cursorial organism is one that is adapted specifically to run. An animal can be considered cursorial if it has the ability to run fast or if it can keep a constant speed for a long distance. "Cursorial" is often used to categorize a certain locomotor mode, which is helpful for biologists who examine behaviors of different animals and the way they move in their environment. Cursorial adaptations can be identified by morphological characteristics, physiological characteristics, maximum speed, and how often running is used in life. There is much debate over how to define a cursorial animal specifically. The most accepted definitions include that a cursorial organism could be considered adapted to long-distance running at high speeds or has the ability to accelerate quickly over short distances. Among vertebrates, animals under 1 kg of mass are rarely considered cursorial, and cursorial behaviors and morphology are thought to only occur at relatively large body masses in mammals. There are a few mammals that have been termed "micro-cursors" that are less than 1 kg in mass and have the ability to run faster than other small animals of similar sizes.
Many vertebrates are limbless, limb-reduced, or apodous, with a body plan consisting of a head and vertebral column, but no adjoining limbs such as legs or fins. Jawless fish are limbless but may have preceded the evolution of vertebrate limbs, whereas numerous reptile and amphibian lineages – and some eels and eel-like fish – independently lost their limbs. Larval amphibians, tadpoles, are also often limbless. No mammals or birds are limbless, but some feature partial limb-loss or limb reduction.

A body plan, Bauplan, or ground plan is a set of morphological features common to many members of a phylum of animals. The vertebrates share one body plan, while invertebrates have many.

Biological or process structuralism is a school of biological thought that objects to an exclusively Darwinian or adaptationist explanation of natural selection such as is described in the 20th century's modern synthesis. It proposes instead that evolution is guided differently, by physical forces which shape the development of an animal's body, and sometimes implies that these forces supersede selection altogether.
Biological constraints are factors which make populations resistant to evolutionary change. One proposed definition of constraint is "A property of a trait that, although possibly adaptive in the environment in which it originally evolved, acts to place limits on the production of new phenotypic variants." Constraint has played an important role in the development of such ideas as homology and body plans.

Carrier's constraint is the observation that air-breathing vertebrates with two lungs that flex their bodies sideways during locomotion find it difficult to move and breathe at the same time, because the sideways flexing expands one lung and compresses the other, shunting stale air from lung to lung instead of expelling it completely to make room for fresh air.

Evolutionary physiology is the study of the biological evolution of physiological structures and processes; that is, the manner in which the functional characteristics of organisms have responded to natural selection or sexual selection or changed by random genetic drift across multiple generations during the history of a population or species. It is a sub-discipline of both physiology and evolutionary biology. Practitioners in the field come from a variety of backgrounds, including physiology, evolutionary biology, ecology, and genetics.
Phylogenetic comparative methods (PCMs) use information on the historical relationships of lineages (phylogenies) to test evolutionary hypotheses. The comparative method has a long history in evolutionary biology; indeed, Charles Darwin used differences and similarities between species as a major source of evidence in The Origin of Species. However, the fact that closely related lineages share many traits and trait combinations as a result of the process of descent with modification means that lineages are not independent. This realization inspired the development of explicitly phylogenetic comparative methods. Initially, these methods were primarily developed to control for phylogenetic history when testing for adaptation; however, in recent years the use of the term has broadened to include any use of phylogenies in statistical tests. Although most studies that employ PCMs focus on extant organisms, many methods can also be applied to extinct taxa and can incorporate information from the fossil record.
The term phylogenetic niche conservatism has seen increasing use in recent years in the scientific literature, though the exact definition has been a matter of some contention. Fundamentally, phylogenetic niche conservatism refers to the tendency of species to retain their ancestral traits. When defined as such, phylogenetic niche conservatism is therefore nearly synonymous with phylogenetic signal. The point of contention is whether or not "conservatism" refers simply to the tendency of species to resemble their ancestors, or implies that "closely related species are more similar than expected based on phylogenetic relationships". If the latter interpretation is employed, then phylogenetic niche conservatism can be seen as an extreme case of phylogenetic signal, and implies that the processes which prevent divergence are in operation in the lineage under consideration. Despite efforts by Jonathan Losos to end this habit, however, the former interpretation appears to frequently motivate scientific research. In this case, phylogenetic niche conservatism might best be considered a form of phylogenetic signal reserved for traits with broad-scale ecological ramifications. Thus, phylogenetic niche conservatism is usually invoked with regards to closely related species occurring in similar environments.


