Related Research Articles
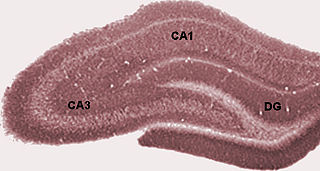
The dentate gyrus (DG) is part of the hippocampal formation in the temporal lobe of the brain, which also includes the hippocampus and the subiculum. The dentate gyrus is part of the hippocampal trisynaptic circuit and is thought to contribute to the formation of new episodic memories, the spontaneous exploration of novel environments and other functions.

Adult neurogenesis is the process in which neurons are generated from neural stem cells in the adult. This process differs from prenatal neurogenesis.

Microglia are a type of neuroglia located throughout the brain and spinal cord. Microglia account for about 10-15% of cells found within the brain. As the resident macrophage cells, they act as the first and main form of active immune defense in the central nervous system (CNS). Microglia originate in the yolk sac under a tightly regulated molecular process. These cells are distributed in large non-overlapping regions throughout the CNS. Microglia are key cells in overall brain maintenance—they are constantly scavenging the CNS for plaques, damaged or unnecessary neurons and synapses, and infectious agents. Since these processes must be efficient to prevent potentially fatal damage, microglia are extremely sensitive to even small pathological changes in the CNS. This sensitivity is achieved in part by the presence of unique potassium channels that respond to even small changes in extracellular potassium. Recent evidence shows that microglia are also key players in the sustainment of normal brain functions under healthy conditions. Microglia also constantly monitor neuronal functions through direct somatic contacts and exert neuroprotective effects when needed.
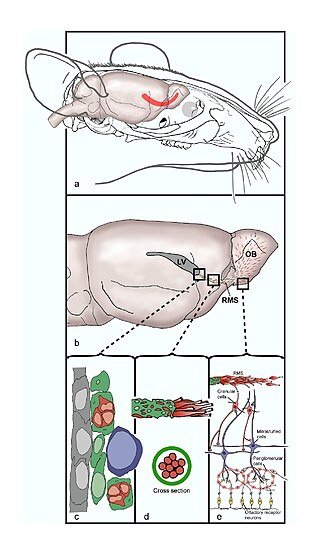
The rostral migratory stream (RMS) is a specialized migratory route found in the brain of some animals along which neuronal precursors that originated in the subventricular zone (SVZ) of the brain migrate to reach the main olfactory bulb (OB). The importance of the RMS lies in its ability to refine and even change an animal's sensitivity to smells, which explains its importance and larger size in the rodent brain as compared to the human brain, as our olfactory sense is not as developed. This pathway has been studied in the rodent, rabbit, and both the squirrel monkey and rhesus monkey. When the neurons reach the OB they differentiate into GABAergic interneurons as they are integrated into either the granule cell layer or periglomerular layer.
HIV-associated neurocognitive disorders (HAND) are neurological disorders associated with HIV infection and AIDS. It is a syndrome of progressive deterioration of memory, cognition, behavior, and motor function in HIV-infected individuals during the late stages of the disease, when immunodeficiency is severe. HAND may include neurological disorders of various severity. HIV-associated neurocognitive disorders are associated with a metabolic encephalopathy induced by HIV infection and fueled by immune activation of macrophages and microglia. These cells are actively infected with HIV and secrete neurotoxins of both host and viral origin. The essential features of HIV-associated dementia (HAD) are disabling cognitive impairment accompanied by motor dysfunction, speech problems and behavioral change. Cognitive impairment is characterised by mental slowness, trouble with memory and poor concentration. Motor symptoms include a loss of fine motor control leading to clumsiness, poor balance and tremors. Behavioral changes may include apathy, lethargy and diminished emotional responses and spontaneity. Histopathologically, it is identified by the infiltration of monocytes and macrophages into the central nervous system (CNS), gliosis, pallor of myelin sheaths, abnormalities of dendritic processes and neuronal loss.

The neuroimmune system is a system of structures and processes involving the biochemical and electrophysiological interactions between the nervous system and immune system which protect neurons from pathogens. It serves to protect neurons against disease by maintaining selectively permeable barriers, mediating neuroinflammation and wound healing in damaged neurons, and mobilizing host defenses against pathogens.
Elizabeth Gould is an American neuroscientist and the Dorman T. Warren Professor of Psychology at Princeton University. She was an early investigator of adult neurogenesis in the hippocampus, a research area that continues to be controversial. In November 2002, Discover magazine listed her as one of the 50 most important women scientists.
Neuroepithelial cells, or neuroectodermal cells, form the wall of the closed neural tube in early embryonic development. The neuroepithelial cells span the thickness of the tube's wall, connecting with the pial surface and with the ventricular or lumenal surface. They are joined at the lumen of the tube by junctional complexes, where they form a pseudostratified layer of epithelium called neuroepithelium.
Neural stem cells (NSCs) are self-renewing, multipotent cells that firstly generate the radial glial progenitor cells that generate the neurons and glia of the nervous system of all animals during embryonic development. Some neural progenitor stem cells persist in highly restricted regions in the adult vertebrate brain and continue to produce neurons throughout life. Differences in the size of the central nervous system are among the most important distinctions between the species and thus mutations in the genes that regulate the size of the neural stem cell compartment are among the most important drivers of vertebrate evolution.

The subventricular zone (SVZ) is a region situated on the outside wall of each lateral ventricle of the vertebrate brain. It is present in both the embryonic and adult brain. In embryonic life, the SVZ refers to a secondary proliferative zone containing neural progenitor cells, which divide to produce neurons in the process of neurogenesis. The primary neural stem cells of the brain and spinal cord, termed radial glial cells, instead reside in the ventricular zone (VZ).

The subgranular zone (SGZ) is a brain region in the hippocampus where adult neurogenesis occurs. The other major site of adult neurogenesis is the subventricular zone (SVZ) in the brain.

Environmental enrichment is the stimulation of the brain by its physical and social surroundings. Brains in richer, more stimulating environments have higher rates of synaptogenesis and more complex dendrite arbors, leading to increased brain activity. This effect takes place primarily during neurodevelopment, but also during adulthood to a lesser degree. With extra synapses there is also increased synapse activity, leading to an increased size and number of glial energy-support cells. Environmental enrichment also enhances capillary vasculation, providing the neurons and glial cells with extra energy. The neuropil expands, thickening the cortex. Research on rodent brains suggests that environmental enrichment may also lead to an increased rate of neurogenesis.
Erythropoietin in neuroprotection is the use of the glycoprotein erythropoietin (Epo) for neuroprotection. Epo controls erythropoiesis, or red blood cell production.
Endogenous regeneration in the brain is the ability of cells to engage in the repair and regeneration process. While the brain has a limited capacity for regeneration, endogenous neural stem cells, as well as numerous pro-regenerative molecules, can participate in replacing and repairing damaged or diseased neurons and glial cells. Another benefit that can be achieved by using endogenous regeneration could be avoiding an immune response from the host.
While researchers have found that moderate alcohol consumption in older adults is associated with better cognition and well-being than abstinence, excessive alcohol consumption is associated with widespread and significant brain lesions. Other data – including investigated brain-scans of 36,678 UK Biobank participants – suggest that even "light" or "moderate" consumption of alcohol by itself harms the brain, such as by reducing brain grey matter volume. This may imply that alternatives and generally aiming for lowest possible consumption could usually be the advisable approach.
Travel outside the Earth's protective atmosphere, magnetosphere, and in free fall can harm human health, and understanding such harm is essential for successful crewed spaceflight. Potential effects on the central nervous system (CNS) are particularly important. A vigorous ground-based cellular and animal model research program will help quantify the risk to the CNS from space radiation exposure on future long distance space missions and promote the development of optimized countermeasures.
Neuroinflammation is inflammation of the nervous tissue. It may be initiated in response to a variety of cues, including infection, traumatic brain injury, toxic metabolites, or autoimmunity. In the central nervous system (CNS), including the brain and spinal cord, microglia are the resident innate immune cells that are activated in response to these cues. The CNS is typically an immunologically privileged site because peripheral immune cells are generally blocked by the blood–brain barrier (BBB), a specialized structure composed of astrocytes and endothelial cells. However, circulating peripheral immune cells may surpass a compromised BBB and encounter neurons and glial cells expressing major histocompatibility complex molecules, perpetuating the immune response. Although the response is initiated to protect the central nervous system from the infectious agent, the effect may be toxic and widespread inflammation as well as further migration of leukocytes through the blood–brain barrier may occur.

Raz Yirmiya is an Israeli behavioral neuroscientist and director of the Laboratory for Psychoneuroimmunology at the Hebrew University of Jerusalem in Israel. He is best known for providing the first experimental evidence for the role of immune system activation in depression, for discovering that disturbances in brain microglia cells underlie some forms of depression, and for elucidating the involvement of inflammatory cytokines in regulation of cognitive and emotional processes.
Neurogenesis is the process by which nervous system cells, the neurons, are produced by neural stem cells (NSCs). In short, it is brain growth in relation to its organization. This occurs in all species of animals except the porifera (sponges) and placozoans. Types of NSCs include neuroepithelial cells (NECs), radial glial cells (RGCs), basal progenitors (BPs), intermediate neuronal precursors (INPs), subventricular zone astrocytes, and subgranular zone radial astrocytes, among others.

Erin M. Gibson is a glial and circadian biologist as well as an assistant professor in the Department of Psychiatry and Behavioral Sciences and the Stanford Center for Sleep Sciences and Medicine at Stanford University. Gibson investigates the role of glial cells in sculpting neural circuits and mechanistically probes how the circadian rhythm modulates glial biology.
References
- 1 2 Bruel-Jungerman, Elodie; Rampon, Claire; Laroche, Serge (2007). "Adult Hippocampal Neurogenesis, Synaptic Plasticity and Memory: Facts and Hypotheses". Reviews in the Neurosciences. 18 (2): 93–114. doi:10.1515/REVNEURO.2007.18.2.93. PMID 17593874. S2CID 21585753.
- ↑ Madsen, T.M; Kristjansen, P.E.G; Bolwig, T.G; Wörtwein, G (2003). "Arrested neuronal proliferation and impaired hippocampal function following fractionated brain irradiation in the adult rat". Neuroscience. 119 (3): 635–42. doi:10.1016/S0306-4522(03)00199-4. PMID 12809684. S2CID 25715150.
- 1 2 Monje, Michelle L.; Palmer, Theo (2003). "Radiation injury and neurogenesis". Current Opinion in Neurology. 16 (2): 129–34. doi:10.1097/00019052-200304000-00002. PMID 12644738.
- ↑ Mendola, Pauline; Selevan, Sherry G.; Gutter, Suzanne; Rice, Deborah (2002). "Environmental factors associated with a spectrum of neurodevelopmental deficits". Mental Retardation and Developmental Disabilities Research Reviews. 8 (3): 188–97. doi:10.1002/mrdd.10033. PMID 12216063.
- 1 2 Hall, P.; Adami, HO; Trichopoulos, D; Pedersen, NL; Lagiou, P; Ekbom, A; Ingvar, M; Lundell, M; Granath, F (2004). "Effect of low doses of ionising radiation in infancy on cognitive function in adulthood: Swedish population based cohort study". BMJ. 328 (7430): 19. doi:10.1136/bmj.328.7430.19. PMC 313898 . PMID 14703539.
- ↑ Asteriadis, I (2004). "Low radiation doses; are they harmful in infancy?". Hellenic Journal of Nuclear Medicine. 7 (1): 2–4. PMID 16868634.
- ↑ "Late Effects of Treatment for Childhood Cancer". National Cancer Institute. 12 April 2012. Retrieved 7 June 2012.
- 1 2 3 Loganovsky, K (2009). "Do Low Doses of Ionizing Radiation Affect the Human Brain?". Data Science Journal. 8: BR13–35. doi: 10.2481/dsj.BR-04 .
- ↑ Otake, M.; Schull, WJ (1998). "Review: Radiation-related brain damage and growth retardation among the prenatally exposed atomic bomb survivors". International Journal of Radiation Biology. 74 (2): 159–71. doi:10.1080/095530098141555. PMID 9712546.
- ↑ NUCLEAR EVENTS AND THEIR CONSEQUENCES by the Borden institute. Chapter 1
- ↑ NUCLEAR EVENTS AND THEIR CONSEQUENCES by the Borden institute. Chapter 7 BEHAVIORAL AND NEUROPHYSIOLOGICAL CHANGES WITH EXPOSURE TO IONIZING RADIATION
- ↑ Mayo Clinic Staff (May 9, 2008). "Symptoms". Radiation sickness. Mayo Clinic. Retrieved 2011-12-24.
- ↑ Picano, Eugenio; Vano, Eliseo; Domenici, Luciano; Bottai, Matteo; Thierry-Chef, Isabelle (2012). "Cancer and non-cancer brain and eye effects of chronic low-dose ionizing radiation exposure". BMC Cancer. 12 (1): 157. doi: 10.1186/1471-2407-12-157 . PMC 3495891 . PMID 22540409.
- ↑ Jain, Prasoon; Mehta, Atul C. (2011). "Radiation terminology". In Wang, Ko-Pen; Mehta, Atul C.; Turner, J. Francis (eds.). Flexible Bronchoscopy. John Wiley & Sons. p. 21. ISBN 978-1-4443-4640-4.
For X rays, the radiation-weighting factor is equal to one; so the equivalent dose in Sv units is equal to the absorbed dose in Gy.
- 1 2 3 4 Monje, Michelle L.; Mizumatsu, Shinichiro; Fike, John R.; Palmer, Theo D. (2002). "Irradiation induces neural precursor-cell dysfunction". Nature Medicine. 8 (9): 955–62. doi:10.1038/nm749. PMID 12161748. S2CID 10347561.
- 1 2 Mizumatsu, S; Monje, ML; Morhardt, DR; Rola, R; Palmer, TD; Fike, JR (2003). "Extreme sensitivity of adult neurogenesis to low doses of X-irradiation". Cancer Research. 63 (14): 4021–7. PMID 12874001.
- 1 2 3 4 Monje, M. L.; Toda, H; Palmer, TD (2003). "Inflammatory Blockade Restores Adult Hippocampal Neurogenesis". Science. 302 (5651): 1760–5. Bibcode:2003Sci...302.1760M. doi: 10.1126/science.1088417 . PMID 14615545. S2CID 36806485.
- ↑ Palmer, Theo D.; Willhoite, Andrew R.; Gage, Fred H. (2000). "Vascular niche for adult hippocampal neurogenesis". The Journal of Comparative Neurology. 425 (4): 479–94. doi:10.1002/1096-9861(20001002)425:4<479::AID-CNE2>3.0.CO;2-3. PMID 10975875. S2CID 46440541.
- 1 2 Louissaint Jr., Abner; Rao, Sudha; Leventhal, Caroline; Goldman, Steven A. (2002). "Coordinated Interaction of Neurogenesis and Angiogenesis in the Adult Songbird Brain". Neuron. 34 (6): 945–60. doi: 10.1016/S0896-6273(02)00722-5 . PMID 12086642.
- ↑ Kanzawa, T; Iwado, E; Aoki, H; Iwamaru, A; Hollingsworth, E F; Sawaya, R; Kondo, S; Kondo, Y (2006). "Ionizing radiation induces apoptosis and inhibits neuronal differentiation in rat neural stem cells via the c-Jun NH2-terminal kinase (JNK) pathway". Oncogene. 25 (26): 3638–48. doi: 10.1038/sj.onc.1209414 . PMID 16491125.
- ↑ Fan, Yang; Liu, Zhengyan; Weinstein, Philip R.; Fike, John R.; Liu, Jialing (2007). "Environmental enrichment enhances neurogenesis and improves functional outcome after cranial irradiation". European Journal of Neuroscience. 25 (1): 38–46. doi:10.1111/j.1460-9568.2006.05269.x. PMID 17241265. S2CID 43259184.