Related Research Articles
Small populations can behave differently from larger populations. They are often the result of population bottlenecks from larger populations, leading to loss of heterozygosity and reduced genetic diversity and loss or fixation of alleles and shifts in allele frequencies. A small population is then more susceptible to demographic and genetic stochastic events, which can impact the long-term survival of the population. Therefore, small populations are often considered at risk of endangerment or extinction, and are often of conservation concern.

In population genetics, gene flow is the transfer of genetic material from one population to another. If the rate of gene flow is high enough, then two populations will have equivalent allele frequencies and therefore can be considered a single effective population. It has been shown that it takes only "one migrant per generation" to prevent populations from diverging due to drift. Populations can diverge due to selection even when they are exchanging alleles, if the selection pressure is strong enough. Gene flow is an important mechanism for transferring genetic diversity among populations. Migrants change the distribution of genetic diversity among populations, by modifying allele frequencies. High rates of gene flow can reduce the genetic differentiation between the two groups, increasing homogeneity. For this reason, gene flow has been thought to constrain speciation and prevent range expansion by combining the gene pools of the groups, thus preventing the development of differences in genetic variation that would have led to differentiation and adaptation. In some cases dispersal resulting in gene flow may also result in the addition of novel genetic variants under positive selection to the gene pool of a species or population

Biological dispersal refers to both the movement of individuals from their birth site to their breeding site, as well as the movement from one breeding site to another . Dispersal is also used to describe the movement of propagules such as seeds and spores. Technically, dispersal is defined as any movement that has the potential to lead to gene flow. The act of dispersal involves three phases: departure, transfer, settlement and there are different fitness costs and benefits associated with each of these phases. Through simply moving from one habitat patch to another, the dispersal of an individual has consequences not only for individual fitness, but also for population dynamics, population genetics, and species distribution. Understanding dispersal and the consequences both for evolutionary strategies at a species level, and for processes at an ecosystem level, requires understanding on the type of dispersal, the dispersal range of a given species, and the dispersal mechanisms involved.
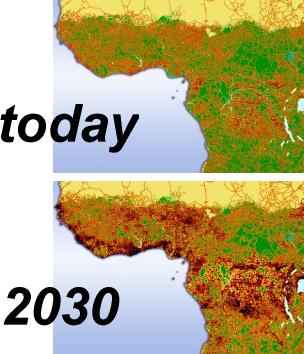
Habitat fragmentation describes the emergence of discontinuities (fragmentation) in an organism's preferred environment (habitat), causing population fragmentation and ecosystem decay. Causes of habitat fragmentation include geological processes that slowly alter the layout of the physical environment, and human activity such as land conversion, which can alter the environment much faster and causes the extinction of many species. More specifically, habitat fragmentation is a process by which large and contiguous habitats get divided into smaller, isolated patches of habitats.

A metapopulation consists of a group of spatially separated populations of the same species which interact at some level. The term metapopulation was coined by Richard Levins in 1969 to describe a model of population dynamics of insect pests in agricultural fields, but the idea has been most broadly applied to species in naturally or artificially fragmented habitats. In Levins' own words, it consists of "a population of populations".

Insular biogeography or island biogeography is a field within biogeography that examines the factors that affect the species richness and diversification of isolated natural communities. The theory was originally developed to explain the pattern of the species–area relationship occurring in oceanic islands. Under either name it is now used in reference to any ecosystem that is isolated due to being surrounded by unlike ecosystems, and has been extended to mountain peaks, seamounts, oases, fragmented forests, and even natural habitats isolated by human land development. The field was started in the 1960s by the ecologists Robert H. MacArthur and E. O. Wilson, who coined the term island biogeography in their inaugural contribution to Princeton's Monograph in Population Biology series, which attempted to predict the number of species that would exist on a newly created island.
The Allee effect is a phenomenon in biology characterized by a correlation between population size or density and the mean individual fitness of a population or species.

Minimum viable population (MVP) is a lower bound on the population of a species, such that it can survive in the wild. This term is commonly used in the fields of biology, ecology, and conservation biology. MVP refers to the smallest possible size at which a biological population can exist without facing extinction from natural disasters or demographic, environmental, or genetic stochasticity. The term "population" is defined as a group of interbreeding individuals in similar geographic area that undergo negligible gene flow with other groups of the species. Typically, MVP is used to refer to a wild population, but can also be used for ex-situ conservation.

Conservation genetics is an interdisciplinary subfield of population genetics that aims to understand the dynamics of genes in a population for the purpose of natural resource management and extinction prevention. Researchers involved in conservation genetics come from a variety of fields including population genetics, natural resources, molecular ecology, biology, evolutionary biology, and systematics. Genetic diversity is one of the three fundamental measures of biodiversity, so it is an important consideration in the wider field of conservation biology.
Genetic load is the difference between the fitness of an average genotype in a population and the fitness of some reference genotype, which may be either the best present in a population, or may be the theoretically optimal genotype. The average individual taken from a population with a low genetic load will generally, when grown in the same conditions, have more surviving offspring than the average individual from a population with a high genetic load. Genetic load can also be seen as reduced fitness at the population level compared to what the population would have if all individuals had the reference high-fitness genotype. High genetic load may put a population in danger of extinction.

Molecular ecology is a field of evolutionary biology that is concerned with applying molecular population genetics, molecular phylogenetics, and more recently genomics to traditional ecological questions. It is virtually synonymous with the field of "Ecological Genetics" as pioneered by Theodosius Dobzhansky, E. B. Ford, Godfrey M. Hewitt, and others. These fields are united in their attempt to study genetic-based questions "out in the field" as opposed to the laboratory. Molecular ecology is related to the field of conservation genetics.
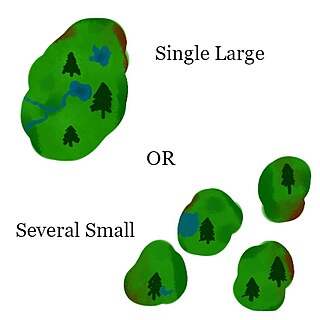
The SLOSS debate was a debate in ecology and conservation biology during the 1970's and 1980's as to whether a single large or several small (SLOSS) reserves were a superior means of conserving biodiversity in a fragmented habitat. Since its inception, multiple alternate theories have been proposed. There have been applications of the concept outside of the original context of habitat conservation.
Inbreeding depression is the reduced biological fitness which has the potential to result from inbreeding. Biological fitness refers to an organism's ability to survive and perpetuate its genetic material. Inbreeding depression is often the result of a population bottleneck. In general, the higher the genetic variation or gene pool within a breeding population, the less likely it is to suffer from inbreeding depression, though inbreeding and outbreeding depression can simultaneously occur.
Extinction vortices are a class of models through which conservation biologists, geneticists and ecologists can understand the dynamics of and categorize extinctions in the context of their causes. This model shows the events that ultimately lead small populations to become increasingly vulnerable as they spiral toward extinction. Developed by M. E. Gilpin and M. E. Soulé in 1986, there are currently four classes of extinction vortices. The first two deal with environmental factors that have an effect on the ecosystem or community level, such as disturbance, pollution, habitat loss etc. Whereas the second two deal with genetic factors such as inbreeding depression and outbreeding depression, genetic drift etc.
Source–sink dynamics is a theoretical model used by ecologists to describe how variation in habitat quality may affect the population growth or decline of organisms.
Extinction threshold is a term used in conservation biology to explain the point at which a species, population or metapopulation, experiences an abrupt change in density or number because of an important parameter, such as habitat loss. It is at this critical value below which a species, population, or metapopulation, will go extinct, though this may take a long time for species just below the critical value, a phenomenon known as extinction debt.
In ecology, the occupancy–abundance (O–A) relationship is the relationship between the abundance of species and the size of their ranges within a region. This relationship is perhaps one of the most well-documented relationships in macroecology, and applies both intra- and interspecifically. In most cases, the O–A relationship is a positive relationship. Although an O–A relationship would be expected, given that a species colonizing a region must pass through the origin and could reach some theoretical maximum abundance and distribution, the relationship described here is somewhat more substantial, in that observed changes in range are associated with greater-than-proportional changes in abundance. Although this relationship appears to be pervasive, and has important implications for the conservation of endangered species, the mechanism(s) underlying it remain poorly understood
An ecological metacommunity is a set of interacting communities which are linked by the dispersal of multiple, potentially interacting species. The term is derived from the field of community ecology, which is primarily concerned with patterns of species distribution, abundance and interactions. Metacommunity ecology combines the importance of local factors and regional factors to explain patterns of species distributions that happen in different spatial scales.
Evolutionary rescue is a process by which a population—that would have gone extinct in the absence of evolution—persists due to natural selection acting on heritable variation. The term was first used in 1995 by Gomulkiewicz and Holt in the context of a sudden environmental change, but the process was studied long before in the context of continuous environmental change and, especially, drug resistance evolution.
Genetic rescue is seen as a mitigation strategy designed to restore genetic diversity and reduce extinction risks in small, isolated and frequently inbred populations. It is largely implemented through translocation, a type of demographic rescue and technical migration that adds individuals to a population to prevent its potential extinction. This demographic rescue may be similar to genetic rescue, as each increase population size and/or fitness. This overlap in meaning has led some researchers to consider a more detailed definition for each type of rescue that details 'assessment and documentation of pre- and post-translocation genetic ancestry'. Not every example of genetic rescue is clearly successful and the current definition of genetic rescue does not mandate that the process result in a 'successful' outcome. Despite an ambiguous definition, genetic rescue is viewed positively, with many perceived successes.
References
- 1 2 3 4 5 Brown JH, Kodric-Brown A. 1977 Turnover rates in insular biogeography: effect of immigration on extinction. Ecology 58, 445– 449. (doi:10.2307/ 1935620)
- 1 2 Richards, C. M. (2000). Inbreeding depression and genetic rescue in a plant metapopulation. American Naturalist, 155, 383– 394.
- 1 2 3 4 Eriksson A, Elı´as-Wolff F, Mehlig B, Manica A. 2014 The emergence of the rescue effect from explicit within- and between-patch dynamics in a metapopulation. Proc. R. Soc. B 281: 20133127. http://dx.doi.org/10.1098/rspb.2013.3127
- 1 2 3 4 5 Gonzalez, A. (1998). "Metapopulation Dynamics, Abundance, and Distribution in a Microecosystem". Science. 281 (5385): 2045–2047. Bibcode:1998Sci...281.2045G. doi:10.1126/science.281.5385.2045. PMID 9748167.
- 1 2 Wu J. and Vankat J.L. 1995. Island biogeography: Theory and applications. In: Nierenberg W.A. (ed.), Encyclopedia of Environmental Biology. Academic Press, San Diego, California, USA, pp. 371–379.
- ↑ Pavlacky DC, Possingham HP, Lowe AJ, Prentis PJ, Green DJ, Goldizen AW. 2012 Anthropogenic landscape change promotes asymmetric dispersal and limits regional patch occupancy in a spatially structured bird population. J. Anim. Ecol. 81,
- ↑ Dornier A, Cheptou P-O. 2012 Determinants of extinction in fragmented plant populations: Crepis sancta (asteraceae) in urban environments. Oecologia 169, 703– 712. (doi:10.1007/s00442-011- 2229-0)
- ↑ Lawson CR, Bennie JJ, Thomas CD, Hodgson JA, Wilson RJ. 2012 Local and landscape management of an expanding range margin under climate change. J. Appl. Ecol. 49, 552– 561.
- 1 2 Ricklefs RE, Cox GW. 1972. Taxon cycles in the West Indian avifauna. The American Naturalist 106: 195–219.
- 1 2 Scott Wilson, Ann E. McKellar, Matthew W. Reudink, Peter P. Marra and Laurene M. Ratcliffe, Density-dependent immigration promotes population stability in a long-distance migratory bird, Population Ecology, 10.1007/s10144-017-0582-5, 59, 2, (169-178), (2017).
- ↑ Diamond, M. (1975). THE ISLAND DILEMMA: LESSONS OF MODERN BIOGEOGRAPHIC STUDIES FOR THE DESIGN OF NATURAL RESERVES
- 1 2 3 Macdonald, D. W., & Willis, K. J. (2013). Key Topics in Conservation Biology 2. Hoboken, NJ, Estados Unidos: Wiley.
- ↑ Harding, C.; Mcnamara, M (2002). "A Unifying Framework for Metapopulation Dynamics". The American Naturalist. 160 (2): 173–85. doi:10.1086/341014. PMID 18707484. S2CID 42154.