
Theoretical ecology is the scientific discipline devoted to the study of ecological systems using theoretical methods such as simple conceptual models, mathematical models, computational simulations, and advanced data analysis. Effective models improve understanding of the natural world by revealing how the dynamics of species populations are often based on fundamental biological conditions and processes. Further, the field aims to unify a diverse range of empirical observations by assuming that common, mechanistic processes generate observable phenomena across species and ecological environments. Based on biologically realistic assumptions, theoretical ecologists are able to uncover novel, non-intuitive insights about natural processes. Theoretical results are often verified by empirical and observational studies, revealing the power of theoretical methods in both predicting and understanding the noisy, diverse biological world.

Inbreeding is the production of offspring from the mating or breeding of individuals or organisms that are closely related genetically. By analogy, the term is used in human reproduction, but more commonly refers to the genetic disorders and other consequences that may arise from expression of deleterious recessive traits resulting from incestuous sexual relationships and consanguinity. Animals avoid incest only rarely.
Small populations can behave differently from larger populations. They are often the result of population bottlenecks from larger populations, leading to loss of heterozygosity and reduced genetic diversity and loss or fixation of alleles and shifts in allele frequencies. A small population is then more susceptible to demographic and genetic stochastic events, which can impact the long-term survival of the population. Therefore, small populations are often considered at risk of endangerment or extinction, and are often of conservation concern.
In population ecology and economics, maximum sustainable yield (MSY) is theoretically, the largest yield that can be taken from a species' stock over an indefinite period. Fundamental to the notion of sustainable harvest, the concept of MSY aims to maintain the population size at the point of maximum growth rate by harvesting the individuals that would normally be added to the population, allowing the population to continue to be productive indefinitely. Under the assumption of logistic growth, resource limitation does not constrain individuals' reproductive rates when populations are small, but because there are few individuals, the overall yield is small. At intermediate population densities, also represented by half the carrying capacity, individuals are able to breed to their maximum rate. At this point, called the maximum sustainable yield, there is a surplus of individuals that can be harvested because growth of the population is at its maximum point due to the large number of reproducing individuals. Above this point, density dependent factors increasingly limit breeding until the population reaches carrying capacity. At this point, there are no surplus individuals to be harvested and yield drops to zero. The maximum sustainable yield is usually higher than the optimum sustainable yield and maximum economic yield.

Genetic diversity is the total number of genetic characteristics in the genetic makeup of a species, it ranges widely from the number of species to differences within species and can be attributed to the span of survival for a species. It is distinguished from genetic variability, which describes the tendency of genetic characteristics to vary.
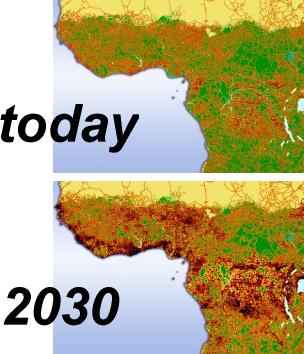
Habitat fragmentation describes the emergence of discontinuities (fragmentation) in an organism's preferred environment (habitat), causing population fragmentation and ecosystem decay. Causes of habitat fragmentation include geological processes that slowly alter the layout of the physical environment, and human activity such as land conversion, which can alter the environment much faster and causes the extinction of many species. More specifically, habitat fragmentation is a process by which large and contiguous habitats get divided into smaller, isolated patches of habitats.

A metapopulation consists of a group of spatially separated populations of the same species which interact at some level. The term metapopulation was coined by Richard Levins in 1969 to describe a model of population dynamics of insect pests in agricultural fields, but the idea has been most broadly applied to species in naturally or artificially fragmented habitats. In Levins' own words, it consists of "a population of populations".
In population genetics and population ecology, population size is a countable quantity representing the number of individual organisms in a population. Population size is directly associated with amount of genetic drift, and is the underlying cause of effects like population bottlenecks and the founder effect. Genetic drift is the major source of decrease of genetic diversity within populations which drives fixation and can potentially lead to speciation events.

Minimum viable population (MVP) is a lower bound on the population of a species, such that it can survive in the wild. This term is commonly used in the fields of biology, ecology, and conservation biology. MVP refers to the smallest possible size at which a biological population can exist without facing extinction from natural disasters or demographic, environmental, or genetic stochasticity. The term "population" is defined as a group of interbreeding individuals in similar geographic area that undergo negligible gene flow with other groups of the species. Typically, MVP is used to refer to a wild population, but can also be used for ex situ conservation.

Conservation genetics is an interdisciplinary subfield of population genetics that aims to understand the dynamics of genes in a population for the purpose of natural resource management, conservation of genetic diversity, and the prevention of species extinction. Scientists involved in conservation genetics come from a variety of fields including population genetics, research in natural resource management, molecular ecology, molecular biology, evolutionary biology, and systematics. The genetic diversity within species is one of the three fundamental components of biodiversity, so it is an important consideration in the wider field of conservation biology.
Genetic load is the difference between the fitness of an average genotype in a population and the fitness of some reference genotype, which may be either the best present in a population, or may be the theoretically optimal genotype. The average individual taken from a population with a low genetic load will generally, when grown in the same conditions, have more surviving offspring than the average individual from a population with a high genetic load. Genetic load can also be seen as reduced fitness at the population level compared to what the population would have if all individuals had the reference high-fitness genotype. High genetic load may put a population in danger of extinction.
Genetic viability is the ability of the genes present to allow a cell, organism or population to survive and reproduce. The term is generally used to mean the chance or ability of a population to avoid the problems of inbreeding. Less commonly genetic viability can also be used in respect to a single cell or on an individual level.

Molecular ecology is a field of evolutionary biology that is concerned with applying molecular population genetics, molecular phylogenetics, and more recently genomics to traditional ecological questions. It is virtually synonymous with the field of "Ecological Genetics" as pioneered by Theodosius Dobzhansky, E. B. Ford, Godfrey M. Hewitt, and others. These fields are united in their attempt to study genetic-based questions "out in the field" as opposed to the laboratory. Molecular ecology is related to the field of conservation genetics.
Inbreeding depression is the reduced biological fitness that has the potential to result from inbreeding. Biological fitness refers to an organism's ability to survive and perpetuate its genetic material. Inbreeding depression is often the result of a population bottleneck. In general, the higher the genetic variation or gene pool within a breeding population, the less likely it is to suffer from inbreeding depression, though inbreeding and outbreeding depression can simultaneously occur.
Extinction vortices are a class of models through which conservation biologists, geneticists and ecologists can understand the dynamics of and categorize extinctions in the context of their causes. This model shows the events that ultimately lead small populations to become increasingly vulnerable as they spiral toward extinction. Developed by M. E. Gilpin and M. E. Soulé in 1986, there are currently four classes of extinction vortices. The first two deal with environmental factors that have an effect on the ecosystem or community level, such as disturbance, pollution, habitat loss etc. Whereas the second two deal with genetic factors such as inbreeding depression and outbreeding depression, genetic drift etc.

A fishery is an area with an associated fish or aquatic population which is harvested for its commercial or recreational value. Fisheries can be wild or farmed. Population dynamics describes the ways in which a given population grows and shrinks over time, as controlled by birth, death, and migration. It is the basis for understanding changing fishery patterns and issues such as habitat destruction, predation and optimal harvesting rates. The population dynamics of fisheries is used by fisheries scientists to determine sustainable yields.
Genetic purging is the reduction of the frequency of a deleterious allele, caused by an increased efficiency of natural selection prompted by inbreeding.

Coexistence theory is a framework to understand how competitor traits can maintain species diversity and stave-off competitive exclusion even among similar species living in ecologically similar environments. Coexistence theory explains the stable coexistence of species as an interaction between two opposing forces: fitness differences between species, which should drive the best-adapted species to exclude others within a particular ecological niche, and stabilizing mechanisms, which maintains diversity via niche differentiation. For many species to be stabilized in a community, population growth must be negative density-dependent, i.e. all participating species have a tendency to increase in density as their populations decline. In such communities, any species that becomes rare will experience positive growth, pushing its population to recover and making local extinction unlikely. As the population of one species declines, individuals of that species tend to compete predominantly with individuals of other species. Thus, the tendency of a population to recover as it declines in density reflects reduced intraspecific competition (within-species) relative to interspecific competition (between-species), the signature of niche differentiation.
Genetic rescue is seen as a mitigation strategy designed to restore genetic diversity and reduce extinction risks in small, isolated and frequently inbred populations. It is largely implemented through translocation, a type of demographic rescue and technical migration that adds individuals to a population to prevent its potential extinction. This demographic rescue may be similar to genetic rescue, as each increase population size and/or fitness. This overlap in meaning has led some researchers to consider a more detailed definition for each type of rescue that details 'assessment and documentation of pre- and post-translocation genetic ancestry'. Not every example of genetic rescue is clearly successful and the current definition of genetic rescue does not mandate that the process result in a 'successful' outcome. Despite an ambiguous definition, genetic rescue is viewed positively, with many perceived successes.
The rescue effect is a phenomenon which was first described by Brown and Kodric-Brown, and is commonly used in metapopulation dynamics and many other disciplines in ecology. This populational process explains how the migration of individuals can increase the persistence of small isolated populations by helping to stabilize a metapopulation, thus reducing the chances of extinction. In other words, immigration can lead to the recolonization of previously extinct patches, promoting the long-term persistence of the network of populations.








