In immunology, autoimmunity is the system of immune responses of an organism against its own healthy cells, tissues and other normal body constituents. Any disease resulting from this type of immune response is termed an "autoimmune disease". Prominent examples include celiac disease, diabetes mellitus type 1, Henoch–Schönlein purpura, systemic lupus erythematosus, Sjögren syndrome, eosinophilic granulomatosis with polyangiitis, Hashimoto's thyroiditis, Graves' disease, idiopathic thrombocytopenic purpura, Addison's disease, rheumatoid arthritis, ankylosing spondylitis, polymyositis, dermatomyositis, and multiple sclerosis. Autoimmune diseases are very often treated with steroids.

The T helper cells (Th cells), also known as CD4+ cells or CD4-positive cells, are a type of T cell that play an important role in the adaptive immune system. They aid the activity of other immune cells by releasing cytokines. They are considered essential in B cell antibody class switching, breaking cross-tolerance in dendritic cells, in the activation and growth of cytotoxic T cells, and in maximizing bactericidal activity of phagocytes such as macrophages and neutrophils. CD4+ cells are mature Th cells that express the surface protein CD4. Genetic variation in regulatory elements expressed by CD4+ cells determines susceptibility to a broad class of autoimmune diseases.
The regulatory T cells (Tregs or Treg cells), formerly known as suppressor T cells, are a subpopulation of T cells that modulate the immune system, maintain tolerance to self-antigens, and prevent autoimmune disease. Treg cells are immunosuppressive and generally suppress or downregulate induction and proliferation of effector T cells. Treg cells express the biomarkers CD4, FOXP3, and CD25 and are thought to be derived from the same lineage as naïve CD4+ cells. Because effector T cells also express CD4 and CD25, Treg cells are very difficult to effectively discern from effector CD4+, making them difficult to study. Research has found that the cytokine transforming growth factor beta (TGF-β) is essential for Treg cells to differentiate from naïve CD4+ cells and is important in maintaining Treg cell homeostasis.

FOXP3, also known as scurfin, is a protein involved in immune system responses. A member of the FOX protein family, FOXP3 appears to function as a master regulator of the regulatory pathway in the development and function of regulatory T cells. Regulatory T cells generally turn the immune response down. In cancer, an excess of regulatory T cell activity can prevent the immune system from destroying cancer cells. In autoimmune disease, a deficiency of regulatory T cell activity can allow other autoimmune cells to attack the body's own tissues.
Co-stimulation is a secondary signal which immune cells rely on to activate an immune response in the presence of an antigen-presenting cell. In the case of T cells, two stimuli are required to fully activate their immune response. During the activation of lymphocytes, co-stimulation is often crucial to the development of an effective immune response. Co-stimulation is required in addition to the antigen-specific signal from their antigen receptors.

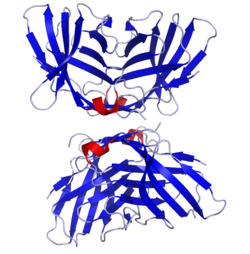
CD28 is one of the proteins expressed on T cells that provide co-stimulatory signals required for T cell activation and survival. T cell stimulation through CD28 in addition to the T-cell receptor (TCR) can provide a potent signal for the production of various interleukins.
Immune tolerance, also known as immunological tolerance or immunotolerance, refers to the immune system's state of unresponsiveness to substances or tissues that would otherwise trigger an immune response. It arises from prior exposure to a specific antigen and contrasts the immune system's conventional role in eliminating foreign antigens. Depending on the site of induction, tolerance is categorized as either central tolerance, occurring in the thymus and bone marrow, or peripheral tolerance, taking place in other tissues and lymph nodes. Although the mechanisms establishing central and peripheral tolerance differ, their outcomes are analogous, ensuring immune system modulation.

The Cluster of differentiation 80 is a B7, type I membrane protein in the immunoglobulin superfamily, with an extracellular immunoglobulin constant-like domain and a variable-like domain required for receptor binding. It is closely related to CD86, another B7 protein (B7-2), and often works in tandem. Both CD80 and CD86 interact with costimulatory receptors CD28, CTLA-4 (CD152) and the p75 neurotrophin receptor.

Cluster of Differentiation 86 is a protein constitutively expressed on dendritic cells, Langerhans cells, macrophages, B-cells, and on other antigen-presenting cells. Along with CD80, CD86 provides costimulatory signals necessary for T cell activation and survival. Depending on the ligand bound, CD86 can signal for self-regulation and cell-cell association, or for attenuation of regulation and cell-cell disassociation.
Abatacept, sold under the brand name Orencia, is a medication used to treat autoimmune diseases like rheumatoid arthritis, by interfering with the immune activity of T cells. It is a modified antibody.
In immunology, peripheral tolerance is the second branch of immunological tolerance, after central tolerance. It takes place in the immune periphery. Its main purpose is to ensure that self-reactive T and B cells which escaped central tolerance do not cause autoimmune disease. Peripheral tolerance can also serve a purpose in preventing an immune response to harmless food antigens and allergens.
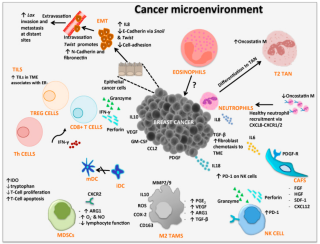
Cancer immunology (immuno-oncology) is an interdisciplinary branch of biology and a sub-discipline of immunology that is concerned with understanding the role of the immune system in the progression and development of cancer; the most well known application is cancer immunotherapy, which utilises the immune system as a treatment for cancer. Cancer immunosurveillance and immunoediting are based on protection against development of tumors in animal systems and (ii) identification of targets for immune recognition of human cancer.

Programmed death-ligand 1 (PD-L1) also known as cluster of differentiation 274 (CD274) or B7 homolog 1 (B7-H1) is a protein that in humans is encoded by the CD274 gene.
Interleukin 35 (IL-35) is a recently discovered anti-inflammatory cytokine from the IL-12 family. Member of IL-12 family - IL-35 is produced by wide range of regulatory lymphocytes and plays a role in immune suppression. IL-35 can block the development of Th1 and Th17 cells by limiting early T cell proliferation.

Protein tyrosine phosphatase non-receptor type 22 (PTPN22) is a cytoplasmatic protein encoded by gene PTPN22 and a member of PEST family of protein tyrosine phosphatases. This protein is also called "PEST-domain Enriched Phosphatase" ("PEP") or "Lymphoid phosphatase" ("LYP"). The name LYP is used strictly for the human protein encoded by PTPN22, but the name PEP is used only for its mouse homolog. However, both proteins have similar biological functions and show 70% identity in amino acid sequence. PTPN22 functions as a negative regulator of T cell receptor (TCR) signaling, which maintains homeostasis of T cell compartment.

ICOS ligand is a protein that in humans is encoded by the ICOSLG gene located at chromosome 21. ICOSLG has also been designated as CD275.

Fc receptor-like protein 3 is a protein that in humans is encoded by the FCRL3 gene.

Immune checkpoints are regulators of the immune system. These pathways are crucial for self-tolerance, which prevents the immune system from attacking cells indiscriminately. However, some cancers can protect themselves from attack by stimulating immune checkpoint targets.

LRBA deficiency is a rare genetic disorder of the immune system. This disorder is caused by a mutation in the gene LRBA. LRBA stands for “lipopolysaccharide (LPS)-responsive and beige-like anchor protein”. This condition is characterized by autoimmunity, lymphoproliferation, and immune deficiency. It was first described by Gabriela Lopez-Herrera from University College London in 2012. Investigators in the laboratory of Dr. Michael Lenardo at National Institute of Allergy and Infectious Diseases, the National Institutes of Health and Dr. Michael Jordan at Cincinnati Children’s Hospital Medical Center later described this condition and therapy in 2015.
CD28 family receptors are a group of regulatory cell surface receptors expressed on immune cells. The CD28 family in turn is a subgroup of the immunoglobulin superfamily.