
The parietal lobe is one of the four major lobes of the cerebral cortex in the brain of mammals. The parietal lobe is positioned above the temporal lobe and behind the frontal lobe and central sulcus.
A gamma wave or gamma rhythm is a pattern of neural oscillation in humans with a frequency between 25 and 140 Hz, the 40 Hz point being of particular interest. Gamma rhythms are correlated with large-scale brain network activity and cognitive phenomena such as working memory, attention, and perceptual grouping, and can be increased in amplitude via meditation or neurostimulation. Altered gamma activity has been observed in many mood and cognitive disorders such as Alzheimer's disease, epilepsy, and schizophrenia.

The human secondary somatosensory cortex is a region of cortex in the parietal operculum on the ceiling of the lateral sulcus.

A cortical homunculus is a distorted representation of the human body, based on a neurological "map" of the areas and proportions of the human brain dedicated to processing motor functions, and/ or sensory functions, for different parts of the body. Nerve fibres—conducting somatosensory information from all over the body—terminate in various areas of the parietal lobe in the cerebral cortex, forming a representational map of the body.

Supplementary eye field (SEF) is the name for the anatomical area of the dorsal medial frontal lobe of the primate cerebral cortex that is indirectly involved in the control of saccadic eye movements. Evidence for a supplementary eye field was first shown by Schlag, and Schlag-Rey. Current research strives to explore the SEF's contribution to visual search and its role in visual salience. The SEF constitutes together with the frontal eye fields (FEF), the intraparietal sulcus (IPS), and the superior colliculus (SC) one of the most important brain areas involved in the generation and control of eye movements, particularly in the direction contralateral to their location. Its precise function is not yet fully known. Neural recordings in the SEF show signals related to both vision and saccades somewhat like the frontal eye fields and superior colliculus, but currently most investigators think that the SEF has a special role in high level aspects of saccade control, like complex spatial transformations, learned transformations, and executive cognitive functions.

The premotor cortex is an area of the motor cortex lying within the frontal lobe of the brain just anterior to the primary motor cortex. It occupies part of Brodmann's area 6. It has been studied mainly in primates, including monkeys and humans. The functions of the premotor cortex are diverse and not fully understood. It projects directly to the spinal cord and therefore may play a role in the direct control of behavior, with a relative emphasis on the trunk muscles of the body. It may also play a role in planning movement, in the spatial guidance of movement, in the sensory guidance of movement, in understanding the actions of others, and in using abstract rules to perform specific tasks. Different subregions of the premotor cortex have different properties and presumably emphasize different functions. Nerve signals generated in the premotor cortex cause much more complex patterns of movement than the discrete patterns generated in the primary motor cortex.

The supplementary motor area (SMA) is a part of the motor cortex of primates that contributes to the control of movement. It is located on the midline surface of the hemisphere just in front of the primary motor cortex leg representation. In monkeys the SMA contains a rough map of the body. In humans the body map is not apparent. Neurons in the SMA project directly to the spinal cord and may play a role in the direct control of movement. Possible functions attributed to the SMA include the postural stabilization of the body, the coordination of both sides of the body such as during bimanual action, the control of movements that are internally generated rather than triggered by sensory events, and the control of sequences of movements. All of these proposed functions remain hypotheses. The precise role or roles of the SMA is not yet known.
Beevor's Axiom is the idea that the brain does not know muscles, only movements. In other words, the brain registers the movements that muscles combine to make, not the individual muscles that are making the movements. Hence, this is why one can sign their name with their foot. Beevor's Axiom was coined by Dr. Charles Edward Beevor, an English neurologist.
A topographic map is the ordered projection of a sensory surface, like the retina or the skin, or an effector system, like the musculature, to one or more structures of the central nervous system. Topographic maps can be found in all sensory systems and in many motor systems.
Premovement neuronal activity in neurophysiological literature refers to neuronal modulations that alter the rate at which neurons fire before a subject produces movement. Through experimentation with multiple animals, predominantly monkeys, it has been shown that several regions of the brain are particularly active and involved in initiation and preparation of movement. Two specific membrane potentials, the bereitschaftspotential, or the BP, and contingent negative variation, or the CNV, play a pivotal role in premovement neuronal activity. Both have been shown to be directly involved in planning and initiating movement. Multiple factors are involved with premovement neuronal activity including motor preparation, inhibition of motor response, programming of the target of movement, closed-looped and open-looped tasks, instructed delay periods, short-lead and long-lead changes, and mirror motor neurons.
Central facial palsy is a symptom or finding characterized by paralysis or paresis of the lower half of one side of the face. It usually results from damage to upper motor neurons of the facial nerve.
The posterior parietal cortex plays an important role in planned movements, spatial reasoning, and attention.

Electrical brain stimulation (EBS), also referred to as focal brain stimulation (FBS), is a form of electrotherapy used as a technique in research and clinical neurobiology to stimulate a neuron or neural network in the brain through the direct or indirect excitation of its cell membrane by using an electric current. EBS is used for research or for therapeutic purposes.

Vittorio Gallese is professor of Psychobiology at the University of Parma, Italy, and was professor in Experimental Aesthetics at the University of London, UK (2016–2018). He is an expert in neurophysiology, cognitive neuroscience, social neuroscience, and philosophy of mind. Gallese is one of the discoverers of mirror neurons. His research attempts to elucidate the functional organization of brain mechanisms underlying social cognition, including action understanding, empathy, language, mindreading and aesthetic experience.
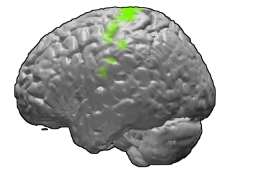
The primary motor cortex is a brain region that in humans is located in the dorsal portion of the frontal lobe. It is the primary region of the motor system and works in association with other motor areas including premotor cortex, the supplementary motor area, posterior parietal cortex, and several subcortical brain regions, to plan and execute voluntary movements. Primary motor cortex is defined anatomically as the region of cortex that contains large neurons known as Betz cells, which, along with other cortical neurons, send long axons down the spinal cord to synapse onto the interneuron circuitry of the spinal cord and also directly onto the alpha motor neurons in the spinal cord which connect to the muscles.
The neuroscience of music is the scientific study of brain-based mechanisms involved in the cognitive processes underlying music. These behaviours include music listening, performing, composing, reading, writing, and ancillary activities. It also is increasingly concerned with the brain basis for musical aesthetics and musical emotion. Scientists working in this field may have training in cognitive neuroscience, neurology, neuroanatomy, psychology, music theory, computer science, and other relevant fields.
Michael Steven Anthony Graziano is an American scientist and novelist who is currently a professor of Psychology and Neuroscience at Princeton University. His scientific research focuses on the brain basis of awareness. He has proposed the "attention schema" theory, an explanation of how, and for what adaptive advantage, brains attribute the property of awareness to themselves. His previous work focused on how the cerebral cortex monitors the space around the body and controls movement within that space. Notably he has suggested that the classical map of the body in motor cortex, the homunculus, is not correct and is better described as a map of complex actions that make up the behavioral repertoire. His publications on this topic have had a widespread impact among neuroscientists but have also generated controversy. His novels rely partly on his background in psychology and are known for surrealism or magic realism. Graziano also composes music including symphonies and string quartets.
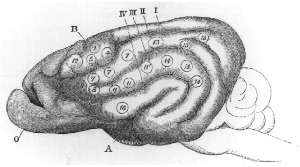
Cortical remapping, also referred to as cortical reorganization, is the process by which an existing cortical map is affected by a stimulus resulting in the creating of a 'new' cortical map. Every part of the body is connected to a corresponding area in the brain which creates a cortical map. When something happens to disrupt the cortical maps such as an amputation or a change in neuronal characteristics, the map is no longer relevant. The part of the brain that is in charge of the amputated limb or neuronal change will be dominated by adjacent cortical regions that are still receiving input, thus creating a remapped area. Remapping can occur in the sensory or motor system. The mechanism for each system may be quite different. Cortical remapping in the somatosensory system happens when there has been a decrease in sensory input to the brain due to deafferentation or amputation, as well as a sensory input increase to an area of the brain. Motor system remapping receives more limited feedback that can be difficult to interpret.
Social cognitive neuroscience is the scientific study of the biological processes underpinning social cognition. Specifically, it uses the tools of neuroscience to study "the mental mechanisms that create, frame, regulate, and respond to our experience of the social world". Social cognitive neuroscience uses the epistemological foundations of cognitive neuroscience, and is closely related to social neuroscience. Social cognitive neuroscience employs human neuroimaging, typically using functional magnetic resonance imaging (fMRI). Human brain stimulation techniques such as transcranial magnetic stimulation and transcranial direct-current stimulation are also used. In nonhuman animals, direct electrophysiological recordings and electrical stimulation of single cells and neuronal populations are utilized for investigating lower-level social cognitive processes.

Eberhard Erich Fetz is an American neuroscientist, academic and researcher. He is a Professor of Physiology and Biophysics and DXARTS at the University of Washington.




