
The retina is the innermost, light-sensitive layer of tissue of the eye of most vertebrates and some molluscs. The optics of the eye create a focused two-dimensional image of the visual world on the retina, which then processes that image within the retina and sends nerve impulses along the optic nerve to the visual cortex to create visual perception. The retina serves a function which is in many ways analogous to that of the film or image sensor in a camera.

A photoreceptor cell is a specialized type of neuroepithelial cell found in the retina that is capable of visual phototransduction. The great biological importance of photoreceptors is that they convert light into signals that can stimulate biological processes. To be more specific, photoreceptor proteins in the cell absorb photons, triggering a change in the cell's membrane potential.
An inhibitory postsynaptic potential (IPSP) is a kind of synaptic potential that makes a postsynaptic neuron less likely to generate an action potential. IPSPs were first investigated in motorneurons by David P. C. Lloyd, John Eccles and Rodolfo Llinás in the 1950s and 1960s. The opposite of an inhibitory postsynaptic potential is an excitatory postsynaptic potential (EPSP), which is a synaptic potential that makes a postsynaptic neuron more likely to generate an action potential. IPSPs can take place at all chemical synapses, which use the secretion of neurotransmitters to create cell to cell signalling. Inhibitory presynaptic neurons release neurotransmitters that then bind to the postsynaptic receptors; this induces a change in the permeability of the postsynaptic neuronal membrane to particular ions. An electric current that changes the postsynaptic membrane potential to create a more negative postsynaptic potential is generated, i.e. the postsynaptic membrane potential becomes more negative than the resting membrane potential, and this is called hyperpolarisation. To generate an action potential, the postsynaptic membrane must depolarize—the membrane potential must reach a voltage threshold more positive than the resting membrane potential. Therefore, hyperpolarisation of the postsynaptic membrane makes it less likely for depolarisation to sufficiently occur to generate an action potential in the postsynaptic neurone.

Rod cells are photoreceptor cells in the retina of the eye that can function in lower light better than the other type of visual photoreceptor, cone cells. Rods are usually found concentrated at the outer edges of the retina and are used in peripheral vision. On average, there are approximately 92 million rod cells in the human retina. Rod cells are more sensitive than cone cells and are almost entirely responsible for night vision. However, rods have little role in color vision, which is the main reason why colors are much less apparent in dim light.

A retinal ganglion cell (RGC) is a type of neuron located near the inner surface of the retina of the eye. It receives visual information from photoreceptors via two intermediate neuron types: bipolar cells and retina amacrine cells. Retina amacrine cells, particularly narrow field cells, are important for creating functional subunits within the ganglion cell layer and making it so that ganglion cells can observe a small dot moving a small distance. Retinal ganglion cells collectively transmit image-forming and non-image forming visual information from the retina in the form of action potential to several regions in the thalamus, hypothalamus, and mesencephalon, or midbrain.

Sensory neurons, also known as afferent neurons, are neurons in the nervous system, that convert a specific type of stimulus, via their receptors, into action potentials or graded receptor potentials. This process is called sensory transduction. The cell bodies of the sensory neurons are located in the dorsal ganglia of the spinal cord.
The receptive field, or sensory space, is a delimited medium where some physiological stimuli can evoke a sensory neuronal response in specific organisms.
As a part of the retina, bipolar cells exist between photoreceptors and ganglion cells. They act, directly or indirectly, to transmit signals from the photoreceptors to the ganglion cells.

Motion perception is the process of inferring the speed and direction of elements in a scene based on visual, vestibular and proprioceptive inputs. Although this process appears straightforward to most observers, it has proven to be a difficult problem from a computational perspective, and difficult to explain in terms of neural processing.

Electroretinography measures the electrical responses of various cell types in the retina, including the photoreceptors, inner retinal cells, and the ganglion cells. Electrodes are placed on the surface of the cornea or on the skin beneath the eye to measure retinal responses. Retinal pigment epithelium (RPE) responses are measured with an EOG test with skin-contact electrodes placed near the canthi. During a recording, the patient's eyes are exposed to standardized stimuli and the resulting signal is displayed showing the time course of the signal's amplitude (voltage). Signals are very small, and typically are measured in microvolts or nanovolts. The ERG is composed of electrical potentials contributed by different cell types within the retina, and the stimulus conditions can elicit stronger response from certain components.
Visual phototransduction is the sensory transduction process of the visual system by which light is detected to yield nerve impulses in the rod cells and cone cells in the retina of the eye in humans and other vertebrates. It relies on the visual cycle, a sequence of biochemical reactions in which a molecule of retinal bound to opsin undergoes photoisomerization, initiates a cascade that signals detection of the photon, and is indirectly restored to its photosensitive isomer for reuse. Phototransduction in some invertebrates such as fruit flies relies on similar processes.
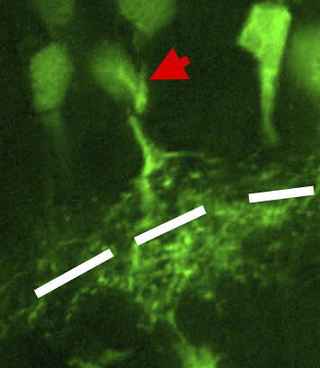
In the anatomy of the eye, amacrine cells are interneurons in the retina. They are named from Greek a– 'non', makr– 'long', and in– 'fiber', because of their short neuronal processes. Amacrine cells are inhibitory neurons, and they project their dendritic arbors onto the inner plexiform layer (IPL), they interact with retinal ganglion cells, and bipolar cells or both of these.
Intrinsically photosensitive retinal ganglion cells (ipRGCs), also called photosensitive retinal ganglion cells (pRGC), or melanopsin-containing retinal ganglion cells (mRGCs), are a type of neuron in the retina of the mammalian eye. The presence of ipRGCs was first suspected in 1927 when rodless, coneless mice still responded to a light stimulus through pupil constriction, This implied that rods and cones are not the only light-sensitive neurons in the retina. Yet research on these cells did not advance until the 1980s. Recent research has shown that these retinal ganglion cells, unlike other retinal ganglion cells, are intrinsically photosensitive due to the presence of melanopsin, a light-sensitive protein. Therefore, they constitute a third class of photoreceptors, in addition to rod and cone cells.

Congenital stationary night blindness (CSNB) is a rare non-progressive retinal disorder. People with CSNB often have difficulty adapting to low light situations due to impaired photoreceptor transmission. These patients may also have reduced visual acuity, myopia, nystagmus, and strabismus. CSNB has two forms -- complete, also known as type-1 (CSNB1), and incomplete, also known as type-2 (CSNB2), which are distinguished by the involvement of different retinal pathways. In CSNB1, downstream neurons called bipolar cells are unable to detect neurotransmission from photoreceptor cells. CSNB1 can be caused by mutations in various genes involved in neurotransmitter detection, including NYX. In CSNB2, the photoreceptors themselves have impaired neurotransmission function; this is caused primarily by mutations in the gene CACNA1F, which encodes a voltage-gated calcium channel important for neurotransmitter release. CSNB has been identified in horses and dogs as the result of mutations in TRPM1, GRM6, and LRIT3 .

A bipolar neuron, or bipolar cell, is a type of neuron that has two extensions. Many bipolar cells are specialized sensory neurons for the transmission of sense. As such, they are part of the sensory pathways for smell, sight, taste, hearing, touch, balance and proprioception. The other shape classifications of neurons include unipolar, pseudounipolar and multipolar. During embryonic development, pseudounipolar neurons begin as bipolar in shape but become pseudounipolar as they mature.

Glutamate receptor, metabotropic 6, also known as GRM6 or mGluR6, is a protein which in humans is encoded by the GRM6 gene.
The ribbon synapse is a type of neuronal synapse characterized by the presence of an electron-dense structure, the synaptic ribbon, that holds vesicles close to the active zone. It is characterized by a tight vesicle-calcium channel coupling that promotes rapid neurotransmitter release and sustained signal transmission. Ribbon synapses undergo a cycle of exocytosis and endocytosis in response to graded changes of membrane potential. It has been proposed that most ribbon synapses undergo a special type of exocytosis based on coordinated multivesicular release. This interpretation has recently been questioned at the inner hair cell ribbon synapse, where it has been instead proposed that exocytosis is described by uniquantal release shaped by a flickering vesicle fusion pore.
Retinal waves are spontaneous bursts of action potentials that propagate in a wave-like fashion across the developing retina. These waves occur before rod and cone maturation and before vision can occur. The signals from retinal waves drive the activity in the dorsal lateral geniculate nucleus (dLGN) and the primary visual cortex. The waves are thought to propagate across neighboring cells in random directions determined by periods of refractoriness that follow the initial depolarization. Retinal waves are thought to have properties that define early connectivity of circuits and synapses between cells in the retina. There is still much debate about the exact role of retinal waves. Some contend that the waves are instructional in the formation of retinogeniculate pathways, while others argue that the activity is necessary but not instructional in the formation of retinogeniculate pathways.
Frank Werblin is Professor of the Graduate School, Division of Neurobiology at the University of California, Berkeley.
AII amacrine cells are a subtype of amacrine cells present in the retina of mammals. AII amacrine cell serve the critical role of transferring light signals from rod photoreceptors to the retinal ganglion cells


