
Nitrification is the biological oxidation of ammonia to nitrate via the intermediary nitrite. Nitrification is an important step in the nitrogen cycle in soil. The process of complete nitrification may occur through separate organisms or entirely within one organism, as in comammox bacteria. The transformation of ammonia to nitrite is usually the rate limiting step of nitrification. Nitrification is an aerobic process performed by small groups of autotrophic bacteria and archaea.
Anaerobic respiration is respiration using electron acceptors other than molecular oxygen (O2). Although oxygen is not the final electron acceptor, the process still uses a respiratory electron transport chain.
Methanogens are anaerobic archaea that produce methane as a byproduct of their energy metabolism, i.e., catabolism. Methane production, or methanogenesis, is the only biochemical pathway for ATP generation in methanogens. All known methanogens belong exclusively to the domain Archaea, although some bacteria, plants, and animal cells are also known to produce methane. However, the biochemical pathway for methane production in these organisms differs from that in methanogens and does not contribute to ATP formation. Methanogens belong to various phyla within the domain Archaea. Previous studies placed all known methanogens into the superphylum Euryarchaeota. However, recent phylogenomic data have led to their reclassification into several different phyla. Methanogens are common in various anoxic environments, such as marine and freshwater sediments, wetlands, the digestive tracts of animals, wastewater treatment plants, rice paddy soil, and landfills. While some methanogens are extremophiles, such as Methanopyrus kandleri, which grows between 84 and 110°C, or Methanonatronarchaeum thermophilum, which grows at a pH range of 8.2 to 10.2 and a Na+ concentration of 3 to 4.8 M, most of the isolates are mesophilic and grow around neutral pH.
Methanogenesis or biomethanation is the formation of methane coupled to energy conservation by microbes known as methanogens. It is the fourth and final stage of anaerobic digestion. Organisms capable of producing methane for energy conservation have been identified only from the domain Archaea, a group phylogenetically distinct from both eukaryotes and bacteria, although many live in close association with anaerobic bacteria. The production of methane is an important and widespread form of microbial metabolism. In anoxic environments, it is the final step in the decomposition of biomass. Methanogenesis is responsible for significant amounts of natural gas accumulations, the remainder being thermogenic.

Anammox, an abbreviation for "anaerobic ammonium oxidation", is a globally important microbial process of the nitrogen cycle that takes place in many natural environments. The bacteria mediating this process were identified in 1999, and were a great surprise for the scientific community. In the anammox reaction, nitrite and ammonium ions are converted directly into diatomic nitrogen and water.
Methanotrophs are prokaryotes that metabolize methane as their source of carbon and chemical energy. They are bacteria or archaea, can grow aerobically or anaerobically, and require single-carbon compounds to survive.
Denitrifying bacteria are a diverse group of bacteria that encompass many different phyla. This group of bacteria, together with denitrifying fungi and archaea, is capable of performing denitrification as part of the nitrogen cycle. Denitrification is performed by a variety of denitrifying bacteria that are widely distributed in soils and sediments and that use oxidized nitrogen compounds such as nitrate and nitrite in the absence of oxygen as a terminal electron acceptor. They metabolize nitrogenous compounds using various enzymes, including nitrate reductase (NAR), nitrite reductase (NIR), nitric oxide reductase (NOR) and nitrous oxide reductase (NOS), turning nitrogen oxides back to nitrogen gas or nitrous oxide.
Microbial metabolism is the means by which a microbe obtains the energy and nutrients it needs to live and reproduce. Microbes use many different types of metabolic strategies and species can often be differentiated from each other based on metabolic characteristics. The specific metabolic properties of a microbe are the major factors in determining that microbe's ecological niche, and often allow for that microbe to be useful in industrial processes or responsible for biogeochemical cycles.
In biology, syntrophy, syntrophism, or cross-feeding is the cooperative interaction between at least two microbial species to degrade a single substrate. This type of biological interaction typically involves the transfer of one or more metabolic intermediates between two or more metabolically diverse microbial species living in close proximity to each other. Thus, syntrophy can be considered an obligatory interdependency and a mutualistic metabolism between different microbial species, wherein the growth of one partner depends on the nutrients, growth factors, or substrates provided by the other(s).
Nitrifying bacteria are chemolithotrophic organisms that include species of genera such as Nitrosomonas, Nitrosococcus, Nitrobacter, Nitrospina, Nitrospira and Nitrococcus. These bacteria get their energy from the oxidation of inorganic nitrogen compounds. Types include ammonia-oxidizing bacteria (AOB) and nitrite-oxidizing bacteria (NOB). Many species of nitrifying bacteria have complex internal membrane systems that are the location for key enzymes in nitrification: ammonia monooxygenase, hydroxylamine oxidoreductase, and nitrite oxidoreductase.

Methanosarcinales is an order of Archaea in the class Methanomicrobia, phylum Methanobacteriota. The order Methanosarcinales contains both methanogenic and methanotrophic lineages, although the latter have so far no pure culture representatives. Methanotrophic lineages of the order Methanosarcinales were initially abbreviated as ANME to distinguich from aerobic methanotrophic bacteria. Currently, those lineages receive their own names such as Ca. Methanoperedens, Ca. Methanocomedens (ANME-2a), Ca.Methanomarinus (ANME-2b), Ca. Methanogaster (ANME-2c), Ca. Methanovorans (ANME-3). The order contains archaeon with one of the largest genome, Methanosarcina acetivorans C2A, genome size 5,75 Mbp.

Archaea is a domain of organisms. Traditionally, Archaea only included its prokaryotic members, but this since has been found to be paraphyletic, as eukaryotes are now known to have evolved from archaea. Even though the domain Archaea cladistically includes eukaryotes, the term "archaea" in English still generally refers specifically to prokaryotic members of Archaea. Archaea were initially classified as bacteria, receiving the name archaebacteria, but this term has fallen out of use.

Greenhouse gas emissions from wetlands of concern consist primarily of methane and nitrous oxide emissions. Wetlands are the largest natural source of atmospheric methane in the world, and are therefore a major area of concern with respect to climate change. Wetlands account for approximately 20–30% of atmospheric methane through emissions from soils and plants, and contribute an approximate average of 161 Tg of methane to the atmosphere per year.
The sulfate-methane transition zone (SMTZ) is a zone in oceans, lakes, and rivers typically found below the sediment surface in which sulfate and methane coexist. The formation of a SMTZ is driven by the diffusion of sulfate down the sediment column and the diffusion of methane up the sediments. At the SMTZ, their diffusion profiles meet and sulfate and methane react with one another, which allows the SMTZ to harbor a unique microbial community whose main form of metabolism is anaerobic oxidation of methane (AOM). The presence of AOM marks the transition from dissimilatory sulfate reduction to methanogenesis as the main metabolism utilized by organisms.
An oxygen minimum zone (OMZ) is characterized as an oxygen-deficient layer in the world's oceans. Typically found between 200 m to 1500 m deep below regions of high productivity, such as the western coasts of continents. OMZs can be seasonal following the spring-summer upwelling season. Upwelling of nutrient-rich water leads to high productivity and labile organic matter, that is respired by heterotrophs as it sinks down the water column. High respiration rates deplete the oxygen in the water column to concentrations of 2 mg/L or less forming the OMZ. OMZs are expanding, with increasing ocean deoxygenation. Under these oxygen-starved conditions, energy is diverted from higher trophic levels to microbial communities that have evolved to use other biogeochemical species instead of oxygen, these species include nitrate, nitrite, sulphate etc. Several Bacteria and Archea have adapted to live in these environments by using these alternate chemical species and thrive. The most abundant phyla in OMZs are Pseudomonadota, Bacteroidota, Actinomycetota, and Planctomycetota.

The hydrothermal vent microbial community includes all unicellular organisms that live and reproduce in a chemically distinct area around hydrothermal vents. These include organisms in the microbial mat, free floating cells, or bacteria in an endosymbiotic relationship with animals. Chemolithoautotrophic bacteria derive nutrients and energy from the geological activity at Hydrothermal vents to fix carbon into organic forms. Viruses are also a part of the hydrothermal vent microbial community and their influence on the microbial ecology in these ecosystems is a burgeoning field of research.
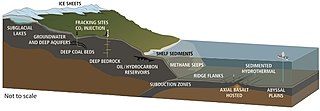
The deep biosphere is the part of the biosphere that resides below the first few meters of the ocean's surface. It extends 10 kilometers below the continental surface and 21 kilometers below the sea surface, at temperatures that may reach beyond 120 °C (248 °F) which is comparable to the maximum temperature where a metabolically active organism has been found. It includes all three domains of life and the genetic diversity rivals that on the surface.

NC10 is a bacterial phylum with candidate status, meaning its members remain uncultured to date. The difficulty in producing lab cultures may be linked to low growth rates and other limiting growth factors.

Candidatus "Methylomirabilis oxyfera" is a candidate species of Gram-negative bacteria belonging to the NC10 phylum, characterized for its capacity to couple anaerobic methane oxidation with nitrite reduction in anoxic environments. To acquire oxygen for methane oxidation, M. oxyfera utilizes an intra-aerobic pathway through the reduction of nitrite (NO2) to dinitrogen (N2) and oxygen.
Methanoperedens nitroreducens is a candidate species of methanotrophic archaea that oxidizes methane by coupling to nitrate reduction.
