In genetics, an operon is a functioning unit of DNA containing a cluster of genes under the control of a single promoter. The genes are transcribed together into an mRNA strand and either translated together in the cytoplasm, or undergo splicing to create monocistronic mRNAs that are translated separately, i.e. several strands of mRNA that each encode a single gene product. The result of this is that the genes contained in the operon are either expressed together or not at all. Several genes must be co-transcribed to define an operon.

Histone methyltransferases (HMT) are histone-modifying enzymes, that catalyze the transfer of one, two, or three methyl groups to lysine and arginine residues of histone proteins. The attachment of methyl groups occurs predominantly at specific lysine or arginine residues on histones H3 and H4. Two major types of histone methyltranferases exist, lysine-specific and arginine-specific. In both types of histone methyltransferases, S-Adenosyl methionine (SAM) serves as a cofactor and methyl donor group.
The genomic DNA of eukaryotes associates with histones to form chromatin. The level of chromatin compaction depends heavily on histone methylation and other post-translational modifications of histones. Histone methylation is a principal epigenetic modification of chromatin that determines gene expression, genomic stability, stem cell maturation, cell lineage development, genetic imprinting, DNA methylation, and cell mitosis.
In molecular biology and genetics, transcriptional regulation is the means by which a cell regulates the conversion of DNA to RNA (transcription), thereby orchestrating gene activity. A single gene can be regulated in a range of ways, from altering the number of copies of RNA that are transcribed, to the temporal control of when the gene is transcribed. This control allows the cell or organism to respond to a variety of intra- and extracellular signals and thus mount a response. Some examples of this include producing the mRNA that encode enzymes to adapt to a change in a food source, producing the gene products involved in cell cycle specific activities, and producing the gene products responsible for cellular differentiation in multicellular eukaryotes, as studied in evolutionary developmental biology.
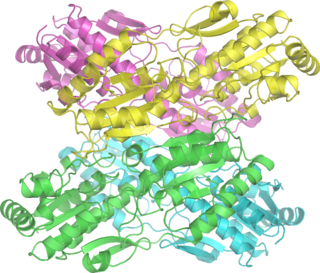
Phosphofructokinase-1 (PFK-1) is one of the most important regulatory enzymes of glycolysis. It is an allosteric enzyme made of 4 subunits and controlled by many activators and inhibitors. PFK-1 catalyzes the important "committed" step of glycolysis, the conversion of fructose 6-phosphate and ATP to fructose 1,6-bisphosphate and ADP. Glycolysis is the foundation for respiration, both anaerobic and aerobic. Because phosphofructokinase (PFK) catalyzes the ATP-dependent phosphorylation to convert fructose-6-phosphate into fructose 1,6-bisphosphate and ADP, it is one of the key regulatory steps of glycolysis. PFK is able to regulate glycolysis through allosteric inhibition, and in this way, the cell can increase or decrease the rate of glycolysis in response to the cell's energy requirements. For example, a high ratio of ATP to ADP will inhibit PFK and glycolysis. The key difference between the regulation of PFK in eukaryotes and prokaryotes is that in eukaryotes PFK is activated by fructose 2,6-bisphosphate. The purpose of fructose 2,6-bisphosphate is to supersede ATP inhibition, thus allowing eukaryotes to have greater sensitivity to regulation by hormones like glucagon and insulin.

Ku is a dimeric protein complex that binds to DNA double-strand break ends and is required for the non-homologous end joining (NHEJ) pathway of DNA repair. Ku is evolutionarily conserved from bacteria to humans. The ancestral bacterial Ku is a homodimer. Eukaryotic Ku is a heterodimer of two polypeptides, Ku70 (XRCC6) and Ku80 (XRCC5), so named because the molecular weight of the human Ku proteins is around 70 kDa and 80 kDa. The two Ku subunits form a basket-shaped structure that threads onto the DNA end. Once bound, Ku can slide down the DNA strand, allowing more Ku molecules to thread onto the end. In higher eukaryotes, Ku forms a complex with the DNA-dependent protein kinase catalytic subunit (DNA-PKcs) to form the full DNA-dependent protein kinase, DNA-PK. Ku is thought to function as a molecular scaffold to which other proteins involved in NHEJ can bind, orienting the double-strand break for ligation.
A bradytroph is a strain of an organism that exhibits slow growth in the absence of an external source of a particular metabolite. This is usually due to a defect in an enzyme required in the metabolic pathway producing this chemical. Such defects are the result of mutations in the genes encoding these enzymes. As the organism can still produce small amounts of the chemical, the mutation is not lethal. In these bradytroph strains, rapid growth occurs when the chemical is present in the cell's growth media and the missing metabolite can be transported into the cell from the external environment. A bradytroph may also be referred to as a "leaky auxotroph".

6-Phosphogluconate dehydrogenase (6PGD) is an enzyme in the pentose phosphate pathway. It forms ribulose 5-phosphate from 6-phosphogluconate:

Glycerol dehydrogenase (EC 1.1.1.6, also known as NAD+-linked glycerol dehydrogenase, glycerol: NAD+ 2-oxidoreductase, GDH, GlDH, GlyDH) is an enzyme in the oxidoreductase family that utilizes the NAD+ to catalyze the oxidation of glycerol to form glycerone (dihydroxyacetone).

In enzymology, an alanine racemase is an enzyme that catalyzes the chemical reaction

Peptidyl arginine deiminase, type IV, also known as PADI4, is a human protein which in humans is encoded by the PADI4 gene.

Protein phosphorylation is a reversible post-translational modification of proteins in which an amino acid residue is phosphorylated by a protein kinase by the addition of a covalently bound phosphate group. Phosphorylation alters the structural conformation of a protein, causing it to become activated, deactivated, or modifying its function. Approximately 13000 human proteins have sites that are phosphorylated.

Protein-arginine deiminase type-2 is an enzyme that in humans is encoded by the PADI2 gene.
Peptidyl arginine deiminase, type III, also known as PADI3, is a protein which in humans is encoded by the PADI3 gene.
Peptidyl arginine deiminase, type I, also known as PADI1, is a protein which in humans is encoded by the PADI1 gene.
The gal operon is a prokaryotic operon, which encodes enzymes necessary for galactose metabolism. Repression of gene expression for this operon works via binding of repressor molecules to two operators. These repressors dimerize, creating a loop in the DNA. The loop as well as hindrance from the external operator prevent RNA polymerase from binding to the promoter, and thus prevent transcription. Additionally, since the metabolism of galactose in the cell is involved in both anabolic and catabolic pathways, a novel regulatory system using two promoters for differential repression has been identified and characterized within the context of the gal operon.
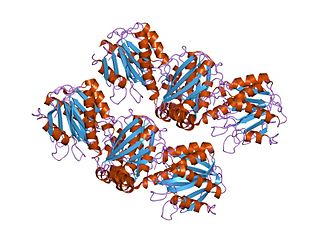
In molecular biology, members of the ArgJ protein family are bifunctional protein that catalyses the first and fifth steps in arginine biosynthesis. The structure has been determined for glutamate N-acetyltransferase 2, an ArgJ-like protein from Streptomyces clavuligerus.

In molecular biology, the LuxR-type DNA-binding HTH domain is a DNA-binding, helix-turn-helix (HTH) domain of about 65 amino acids. It is present in transcription regulators of the LuxR/FixJ family of response regulators. The domain is named after Vibrio fischeri luxR, a transcriptional activator for quorum-sensing control of luminescence. LuxR-type HTH domain proteins occur in a variety of organisms. The DNA-binding HTH domain is usually located in the C-terminal region of the protein; the N-terminal region often containing an autoinducer-binding domain or a response regulatory domain. Most luxR-type regulators act as transcription activators, but some can be repressors or have a dual role for different sites. LuxR-type HTH regulators control a wide variety of activities in various biological processes.
In molecular biology, the SR1 RNA is a small RNA (sRNA) produced by species of Bacillus and closely related bacteria. It is a dual-function RNA which acts both as a protein-coding RNA and as a regulatory sRNA.

Roger Brent is an American biologist known for his work on gene regulation and systems biology. He studies the quantitative behaviors of cell signaling systems and the origins and consequences of variation in them. He is Full Member in the Division of Basic Sciences at the Fred Hutchinson Cancer Research Center and an Affiliate Professor of Genome Sciences at the University of Washington.
Monocercomonoides is a genus of flagellate Excavata belonging to the order Oxymonadida. It was established by Bernard V. Travis and was first described as those with "polymastiginid flagellates having three anterior flagella and a trailing one originating at a single basal granule located in front of the anteriorly positioned nucleus, and a more or less well-defined axostyle". It is the first eukaryotic genus to be found to completely lack mitochondria, and all hallmark proteins responsible for mitochondrial function. Data suggests that the absence of mitochondria is not an ancestral feature, but rather due to secondary loss. Monocercomonoides sp. was found to obtain energy through an enzymatic action of nutrients absorbed from the environment. The genus has replaced the iron-sulphur cluster assembly pathway with a cytosolic sulphur mobilization system, likely acquired by horizontal gene transfer from a eubacterium of a common ancestor of oxymonads. These organisms are significant because they overrule the notion that eukaryotes must have mitochondria to properly function. The genome of Monocercomonoides has approximately 75 million base pairs, with 16629 predicted protein-coding genes.
