
The cell is the basic structural and functional unit of all forms of life. Every cell consists of cytoplasm enclosed within a membrane; many cells contain organelles, each with a specific function. The term comes from the Latin word cellula meaning 'small room'. Most cells are only visible under a microscope. Cells emerged on Earth about 4 billion years ago. All cells are capable of replication, protein synthesis, and motility.
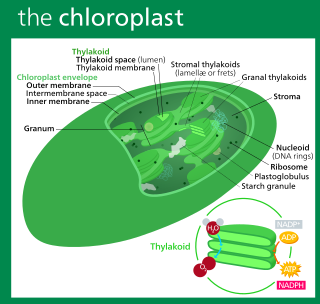
A chloroplast is a type of membrane-bound organelle known as a plastid that conducts photosynthesis mostly in plant and algal cells. The photosynthetic pigment chlorophyll captures the energy from sunlight, converts it, and stores it in the energy-storage molecules ATP and NADPH while freeing oxygen from water in the cells. The ATP and NADPH is then used to make organic molecules from carbon dioxide in a process known as the Calvin cycle. Chloroplasts carry out a number of other functions, including fatty acid synthesis, amino acid synthesis, and the immune response in plants. The number of chloroplasts per cell varies from one, in unicellular algae, up to 100 in plants like Arabidopsis and wheat.
In cell biology, an organelle is a specialized subunit, usually within a cell, that has a specific function. The name organelle comes from the idea that these structures are parts of cells, as organs are to the body, hence organelle, the suffix -elle being a diminutive. Organelles are either separately enclosed within their own lipid bilayers or are spatially distinct functional units without a surrounding lipid bilayer. Although most organelles are functional units within cells, some function units that extend outside of cells are often termed organelles, such as cilia, the flagellum and archaellum, and the trichocyst.
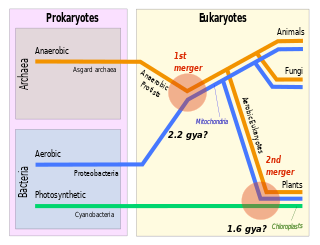
Symbiogenesis is the leading evolutionary theory of the origin of eukaryotic cells from prokaryotic organisms. The theory holds that mitochondria, plastids such as chloroplasts, and possibly other organelles of eukaryotic cells are descended from formerly free-living prokaryotes taken one inside the other in endosymbiosis. Mitochondria appear to be phylogenetically related to Rickettsiales bacteria, while chloroplasts are thought to be related to cyanobacteria.
A plastid, pl. plastids, is a membrane-bound organelle found in the cells of plants, algae, and some other eukaryotic organisms. They are considered to be intracellular endosymbiotic cyanobacteria.
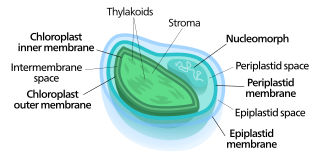
Nucleomorphs are small, vestigial eukaryotic nuclei found between the inner and outer pairs of membranes in certain plastids. They are thought to be vestiges of primitive red and green algal nuclei that were engulfed by a larger eukaryote. Because the nucleomorph lies between two sets of membranes, nucleomorphs support the endosymbiotic theory and are evidence that the plastids containing them are complex plastids. Having two sets of membranes indicate that the plastid, a prokaryote, was engulfed by a eukaryote, an alga, which was then engulfed by another eukaryote, the host cell, making the plastid an example of secondary endosymbiosis.
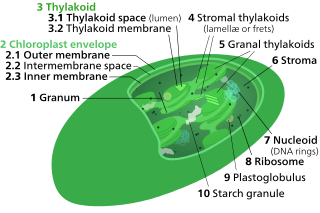
Chloroplasts contain several important membranes, vital for their function. Like mitochondria, chloroplasts have a double-membrane envelope, called the chloroplast envelope, but unlike mitochondria, chloroplasts also have internal membrane structures called thylakoids. Furthermore, one or two additional membranes may enclose chloroplasts in organisms that underwent secondary endosymbiosis, such as the euglenids and chlorarachniophytes.
Gymnophryidae is a small family of amoeboids that lack shells and produce thin, reticulose pseudopods. These contain microtubules and have a granular appearance, owing to the presence of extrusomes, but are distinct from the pseudopods of Foraminifera. They are included among the Cercozoa, but differ from other cercozoans in having mitochondria with flat cristae, rather than tubular cristae.
An apicoplast is a derived non-photosynthetic plastid found in most Apicomplexa, including Toxoplasma gondii, and Plasmodium falciparum and other Plasmodium spp., but not in others such as Cryptosporidium. It originated from algae through secondary endosymbiosis; there is debate as to whether this was a green or red alga. The apicoplast is surrounded by four membranes within the outermost part of the endomembrane system. The apicoplast hosts important metabolic pathways like fatty acid synthesis, isoprenoid precursor synthesis and parts of the heme biosynthetic pathway.

Paulinella is a genus of at least eleven species including both freshwater and marine amoeboids. Like many members of euglyphids it is covered by rows of siliceous scales, and use filose pseudopods to crawl over the substrate of the benthic zone.
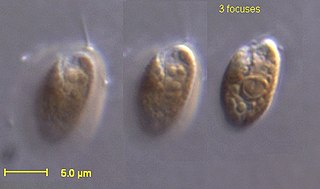
Rhodomonas is a genus of cryptomonads. It is characterized by its red colour, the square-shaped plates of its inner periplast, its short furrow ending in a gullet, and a distinctly shaped chloroplast closely associated with its nucleomorph. Historically, Rhodomonas was characterized by its red chloroplast alone, but this no longer occurs as its taxonomy has become increasingly based on molecular and cellular data. Currently, there is some debate about the taxonomic validity of Rhodomonas as a genus and further research is needed to verify its taxonomic status. Rhodomonas is typically found in marine environments, although freshwater reports exist. It is commonly used as a live feed for various aquaculture species.
Geminigera /ˌdʒɛmɪnɪˈdʒɛɹə/ is a genus of cryptophyte from the family Geminigeraceae. Named for its unique pyrenoids, Geminigera is a genus with a single mixotrophic species. It was discovered in 1968 and is known for living in very cold temperatures such as under the Antarctic ice. While originally considered to be part of the genus Cryptomonas, the genus Geminigera was officially described in 1991 by D. R. A. Hill.

Guillardia is a genus of marine biflagellate cryptomonad algae with a plastid obtained through secondary endosymbiosis of a red alga.

Mesodinium rubrum is a species of ciliates. It constitutes a plankton community and is found throughout the year, most abundantly in spring and fall, in coastal areas. Although discovered in 1908, its scientific importance came into light in the late 1960s when it attracted scientists by the recurrent red colouration it caused by forming massive blooms, that cause red tides in the oceans.

Kraken is a genus of amoebae within the Cercozoa, containing the sole species Kraken carinae. These amoebae are characterized by a small round cell body and a network of thin and very long filopodia that can reach up to a mm in diameter. Kraken amoebae feed on bacteria and live in freshwater and soil systems.
Platysulcus tardus is an eukaryotic microorganism that was recently discovered to be the earliest diverging lineage of the Heterokont phylogenetic tree. It is the only member of the family Platysulcidae, order Platysulcida and class Platysulcea.
Durinskia is a genus of dinoflagellates that can be found in freshwater and marine environments. This genus was created to accommodate its type species, Durinskia baltica, after major classification discrepancies were found. While Durinskia species appear to be typical dinoflagellates that are armored with cellulose plates called theca, the presence of a pennate diatom-derived tertiary endosymbiont is their most defining characteristic. This genus is significant to the study of endosymbiotic events and organelle integration since structures and organelle genomes in the tertiary plastids are not reduced. Like some dinoflagellates, species in Durinskia may cause blooms.

Viridiraptoridae, previously known as clade X, is a clade of heterotrophic protists in the phylum Cercozoa. They're a family of glissomonads, a group containing a vast, mostly undescribed diversity of soil and freshwater organisms.
Orciraptor is a genus of heterotrophic protists, containing the single species Orciraptor agilis. It belongs to the family Viridiraptoridae, in the phylum Cercozoa.
Bihospites is a genus of symbiontid euglenozoans characterized by the presence of two species of epibiotic bacteria on the cell surface. Bihospites cells are clear, biflagellated, and uninucleated, that range between 40–120 μm long and 15–30 μm wide. Bihospites, as well as other members of the symbiontids, are found in semi-anoxic to anoxic sediments in benthic marine environments. Each cell surface is covered by both rod-shaped and spherical-shaped epibiotic bacteria that may share a commensalistic or mutualistic relationship with Bihospites host cells. Bihospites cells are highly contractile and contain several morphological synapomorphies which are present in euglenozoans, however they also contain several unique morphological traits including a unique C-shaped feeding apparatus.
