
Biogeography is the study of the distribution of species and ecosystems in geographic space and through geological time. Organisms and biological communities often vary in a regular fashion along geographic gradients of latitude, elevation, isolation and habitat area. Phytogeography is the branch of biogeography that studies the distribution of plants. Zoogeography is the branch that studies distribution of animals. Mycogeography is the branch that studies distribution of fungi, such as mushrooms.
Phylogeography is the study of the historical processes that may be responsible for the past to present geographic distributions of genealogical lineages. This is accomplished by considering the geographic distribution of individuals in light of genetics, particularly population genetics.
The metabolic theory of ecology (MTE) is the ecological component of the more general Metabolic Scaling Theory and Kleiber's law. It posits that the metabolic rate of organisms is the fundamental biological rate that governs most observed patterns in ecology. MTE is part of a larger set of theory known as metabolic scaling theory that attempts to provide a unified theory for the importance of metabolism in driving pattern and process in biology from the level of cells all the way to the biosphere.

Biological dispersal refers to both the movement of individuals from their birth site to their breeding site, as well as the movement from one breeding site to another . Dispersal is also used to describe the movement of propagules such as seeds and spores. Technically, dispersal is defined as any movement that has the potential to lead to gene flow. The act of dispersal involves three phases: departure, transfer, settlement and there are different fitness costs and benefits associated with each of these phases. Through simply moving from one habitat patch to another, the dispersal of an individual has consequences not only for individual fitness, but also for population dynamics, population genetics, and species distribution. Understanding dispersal and the consequences both for evolutionary strategies at a species level, and for processes at an ecosystem level, requires understanding on the type of dispersal, the dispersal range of a given species, and the dispersal mechanisms involved.

Endemism is the state of a species being found in a single defined geographic location, such as an island, state, nation, country or other defined zone; organisms that are indigenous to a place are not endemic to it if they are also found elsewhere. For example, the Cape sugarbird is found exclusively in southwestern South Africa and is therefore said to be endemic to that particular part of the world.
Macroecology is the subfield of ecology that deals with the study of relationships between organisms and their environment at large spatial scales to characterise and explain statistical patterns of abundance, distribution and diversity. The term was coined in a small monograph published in Spanish in 1971 by Guillermo Sarmiento and Maximina Monasterio, two Venezuelan researchers working in tropical savanna ecosystems and later used by James Brown of the University of New Mexico and Brian Maurer of Michigan State University in a 1989 paper in Science.
Insular biogeography or island biogeography is a field within biogeography that examines the factors that affect the species richness and diversification of isolated natural communities. The theory was originally developed to explain the pattern of the species–area relationship occurring in oceanic islands. Under either name it is now used in reference to any ecosystem that is isolated due to being surrounded by unlike ecosystems, and has been extended to mountain peaks, seamounts, oases, fragmented forests, and even natural habitats isolated by human land development. The field was started in the 1960s by the ecologists Robert H. MacArthur and E. O. Wilson, who coined the term island biogeography in their inaugural contribution to Princeton's Monograph in Population Biology series, which attempted to predict the number of species that would exist on a newly created island.

Kleiber's law, named after Max Kleiber for his biology work in the early 1930s, is the observation that, for the vast majority of animals, an animal's metabolic rate scales to the 3⁄4 power of the animal's mass. Symbolically: if q0 is the animal's metabolic rate, and M is the animal's mass, then Kleiber's law states that q0~M3/4. Thus, over the same time span, a cat having a mass 100 times that of a mouse will consume only about 32 times the energy the mouse uses.
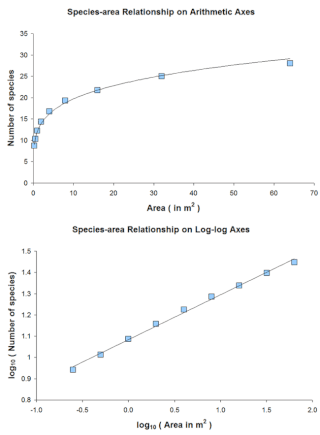
The species-area relationship or species-area curve describes the relationship between the area of a habitat, or of part of a habitat, and the number of species found within that area. Larger areas tend to contain larger numbers of species, and empirically, the relative numbers seem to follow systematic mathematical relationships. The species-area relationship is usually constructed for a single type of organism, such as all vascular plants or all species of a specific trophic level within a particular site. It is rarely if ever, constructed for all types of organisms if simply because of the prodigious data requirements. It is related but not identical to the species discovery curve.

Species richness, or biodiversity, increases from the poles to the tropics for a wide variety of terrestrial and marine organisms, often referred to as the latitudinal diversity gradient. The latitudinal diversity gradient is one of the most widely recognized patterns in ecology. It has been observed to varying degrees in Earth's past. A parallel trend has been found with elevation, though this is less well-studied.
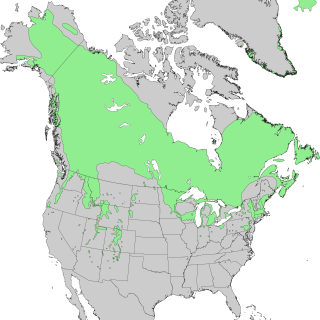
Species distribution —or speciesdispersion — is the manner in which a biological taxon is spatially arranged. The geographic limits of a particular taxon's distribution is its range, often represented as shaded areas on a map. Patterns of distribution change depending on the scale at which they are viewed, from the arrangement of individuals within a small family unit, to patterns within a population, or the distribution of the entire species as a whole (range). Species distribution is not to be confused with dispersal, which is the movement of individuals away from their region of origin or from a population center of high density.
In landscape ecology, landscape connectivity is, broadly, "the degree to which the landscape facilitates or impedes movement among resource patches". Alternatively, connectivity may be a continuous property of the landscape and independent of patches and paths. Connectivity includes both structural connectivity and functional connectivity. Functional connectivity includes actual connectivity and potential connectivity in which movement paths are estimated using the life-history data.
In ecology, the occupancy–abundance (O–A) relationship is the relationship between the abundance of species and the size of their ranges within a region. This relationship is perhaps one of the most well-documented relationships in macroecology, and applies both intra- and interspecifically. In most cases, the O–A relationship is a positive relationship. Although an O–A relationship would be expected, given that a species colonizing a region must pass through the origin and could reach some theoretical maximum abundance and distribution, the relationship described here is somewhat more substantial, in that observed changes in range are associated with greater-than-proportional changes in abundance. Although this relationship appears to be pervasive, and has important implications for the conservation of endangered species, the mechanism(s) underlying it remain poorly understood

The Theory of Island Biogeography is a 1967 book by the ecologist Robert MacArthur and the biologist Edward O. Wilson. It is widely regarded as a seminal piece in island biogeography and ecology. The Princeton University Press reprinted the book in 2001 as a part of the "Princeton Landmarks in Biology" series. The book popularized the theory that insular biota maintain a dynamic equilibrium between immigration and extinction rates. The book also popularized the concepts and terminology of r/K selection theory.
Defaunation is the global, local or functional extinction of animal populations or species from ecological communities. The growth of the human population, combined with advances in harvesting technologies, has led to more intense and efficient exploitation of the environment. This has resulted in the depletion of large vertebrates from ecological communities, creating what has been termed "empty forest". Defaunation differs from extinction; it includes both the disappearance of species and declines in abundance. Defaunation effects were first implied at the Symposium of Plant-Animal Interactions at the University of Campinas, Brazil in 1988 in the context of Neotropical forests. Since then, the term has gained broader usage in conservation biology as a global phenomenon.
A dispersal vector is an agent of biological dispersal that moves a dispersal unit, or organism, away from its birth population to another location or population in which the individual will reproduce. These dispersal units can range from pollen to seeds to fungi to entire organisms.
In macroecology and community ecology, an occupancy frequency distribution (OFD) is the distribution of the numbers of species occupying different numbers of areas. It was first reported in 1918 by the Danish botanist Christen C. Raunkiær in his study on plant communities. The OFD is also known as the species-range size distribution in literature.
Microbial biogeography is a subset of biogeography, a field that concerns the distribution of organisms across space and time. Although biogeography traditionally focused on plants and larger animals, recent studies have broadened this field to include distribution patterns of microorganisms. This extension of biogeography to smaller scales—known as "microbial biogeography"—is enabled by ongoing advances in genetic technologies.
The rescue effect is a phenomenon which was first described by Brown and Kodric-Brown, and is commonly used in metapopulation dynamics and many other disciplines in ecology. This populational process explains how the migration of individuals can increase the persistence of small isolated populations by helping to stabilize a metapopulation, thus reducing the chances of extinction. In other words, immigration can lead to the recolonization of previously extinct patches, promoting the long-term persistence of the network of populations.





