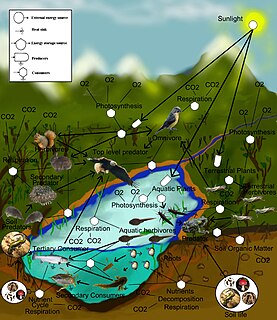
Theoretical ecology is the scientific discipline devoted to the study of ecological systems using theoretical methods such as simple conceptual models, mathematical models, computational simulations, and advanced data analysis. Effective models improve understanding of the natural world by revealing how the dynamics of species populations are often based on fundamental biological conditions and processes. Further, the field aims to unify a diverse range of empirical observations by assuming that common, mechanistic processes generate observable phenomena across species and ecological environments. Based on biologically realistic assumptions, theoretical ecologists are able to uncover novel, non-intuitive insights about natural processes. Theoretical results are often verified by empirical and observational studies, revealing the power of theoretical methods in both predicting and understanding the noisy, diverse biological world.

In ecology, a niche is the match of a species to a specific environmental condition. It describes how an organism or population responds to the distribution of resources and competitors and how it in turn alters those same factors. "The type and number of variables comprising the dimensions of an environmental niche vary from one species to another [and] the relative importance of particular environmental variables for a species may vary according to the geographic and biotic contexts".

In ecology, the competitive exclusion principle, sometimes referred to as Gause's law, is a proposition named for Georgy Gause that two species competing for the same limited resource cannot coexist at constant population values. When one species has even the slightest advantage over another, the one with the advantage will dominate in the long term. This leads either to the extinction of the weaker competitor or to an evolutionary or behavioral shift toward a different ecological niche. The principle has been paraphrased in the maxim "complete competitors can not coexist".

The unified neutral theory of biodiversity and biogeography is a theory and the title of a monograph by ecologist Stephen Hubbell. The hypothesis aims to explain the diversity and relative abundance of species in ecological communities. Like other neutral theories of ecology, Hubbell assumes that the differences between members of an ecological community of trophically similar species are "neutral", or irrelevant to their success. This implies that niche differences do not influence abundance and the abundance of each species follows a random walk. The theory has sparked controversy, and some authors consider it a more complex version of other null models that fit the data better.

The species-area relationship or species-area curve describes the relationship between the area of a habitat, or of part of a habitat, and the number of species found within that area. Larger areas tend to contain larger numbers of species, and empirically, the relative numbers seem to follow systematic mathematical relationships. The species-area relationship is usually constructed for a single type of organism, such as all vascular plants or all species of a specific trophic level within a particular site. It is rarely if ever, constructed for all types of organisms if simply because of the prodigious data requirements. It is related but not identical to the species discovery curve.

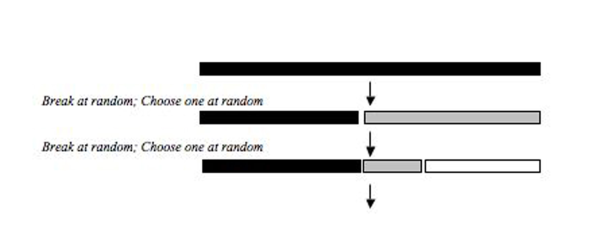
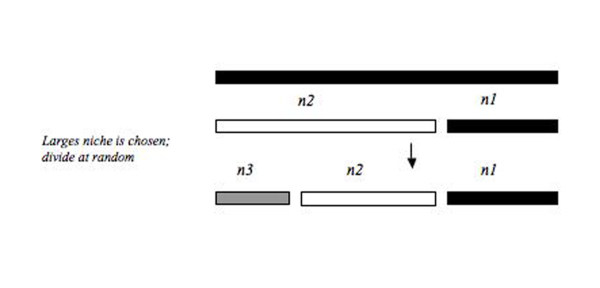
Species use restricted ecological niches, and the niches of all species are segregated, often with much overlap, by the use of different habitats, different geographic areas and seasons, and different food resources, to mention only a few of the many niche dimensions. The causes of niche restriction and segregation are important problems in evolutionary ecology.

In ecology, local abundance is the relative representation of a species in a particular ecosystem. It is usually measured as the number of individuals found per sample. The ratio of abundance of one species to one or multiple other species living in an ecosystem is referred to as relative species abundances. Both indicators are relevant for computing biodiversity.
In ecology, the occupancy–abundance (O–A) relationship is the relationship between the abundance of species and the size of their ranges within a region. This relationship is perhaps one of the most well-documented relationships in macroecology, and applies both intra- and interspecifically. In most cases, the O–A relationship is a positive relationship. Although an O–A relationship would be expected, given that a species colonizing a region must pass through the origin and could reach some theoretical maximum abundance and distribution, the relationship described here is somewhat more substantial, in that observed changes in range are associated with greater-than-proportional changes in abundance. Although this relationship appears to be pervasive, and has important implications for the conservation of endangered species, the mechanism(s) underlying it remain poorly understood
In spatial ecology and macroecology, scaling pattern of occupancy (SPO), also known as the area-of-occupancy (AOO) is the way in which species distribution changes across spatial scales. In physical geography and image analysis, it is similar to the modifiable areal unit problem. Simon A. Levin (1992) states that the problem of relating phenomena across scales is the central problem in biology and in all of science. Understanding the SPO is thus one central theme in ecology.
Community assembly rules are a set of controversial rules in ecology, first proposed by Jared Diamond.

The Theory of Island Biogeography is a 1967 book by the ecologist Robert MacArthur and the biologist Edward O. Wilson. It is widely regarded as a seminal piece in island biogeography and ecology. The Princeton University Press reprinted the book in 2001 as a part of the "Princeton Landmarks in Biology" series. The book popularized the theory that insular biota maintain a dynamic equilibrium between immigration and extinction rates. The book also popularized the concepts and terminology of r/K selection theory.
An ecological metacommunity is a set of interacting communities which are linked by the dispersal of multiple, potentially interacting species. The term is derived from the field of community ecology, which is primarily concerned with patterns of species distribution, abundance and interactions. Metacommunity ecology combines the importance of local factors and regional factors to explain patterns of species distributions that happen in different spatial scales.
Ecological forecasting uses knowledge of physics, ecology and physiology to predict how ecological populations, communities, or ecosystems will change in the future in response to environmental factors such as climate change. The ultimate goal of the approach is to provide people such as resource managers and designers of marine reserves with information that they can then use to respond, in advance, to future changes, a form of adaptation to global warming.

The body size-species richness distribution is a pattern observed in the way taxa are distributed over large spatial scales. The number of species that exhibit small body size generally far exceed the number of species that are large-bodied. Macroecology has long sought to understand the mechanisms that underlie the patterns of biodiversity, such as the body size-species richness pattern.
Relative species abundance is a component of biodiversity and refers to how common or rare a species is relative to other species in a defined location or community. Relative abundance is the percent composition of an organism of a particular kind relative to the total number of organisms in the area. Relative species abundances tend to conform to specific patterns that are among the best-known and most-studied patterns in macroecology. Different populations in a community exist in relative proportions; this idea is known as relative abundance.
Limiting similarity is a concept in theoretical ecology and community ecology that proposes the existence of a maximum level of niche overlap between two given species that will allow continued coexistence.

Species distribution modelling (SDM), also known as environmental(or ecological) niche modelling (ENM), habitat modelling, predictive habitat distribution modelling, and range mapping uses computer algorithms to predict the distribution of a species across geographic space and time using environmental data. The environmental data are most often climate data, but can include other variables such as soil type, water depth, and land cover. SDMs are used in several research areas in conservation biology, ecology and evolution. These models can be used to understand how environmental conditions influence the occurrence or abundance of a species, and for predictive purposes. Predictions from an SDM may be of a species’ future distribution under climate change, a species’ past distribution in order to assess evolutionary relationships, or the potential future distribution of an invasive species. Predictions of current and/or future habitat suitability can be useful for management applications.
No-analog in paleoecology and ecological forecasting are those communities or climates that are compositionally different from a baseline for measurement of communities or climate, which is typically modern. Alternative naming conventions to describe no-analog communities and climates may include novel, emerging, mosaic, disharmonious and intermingled.
In macroecology and community ecology, an occupancy frequency distribution (OFD) is the distribution of the numbers of species occupying different numbers of areas. It was first reported in 1918 by the Danish botanist Christen C. Raunkiær in his study on plant communities. The OFD is also known as the species-range size distribution in literature.
The ecological and biogeographical concept of the species pool describes all species available that could potentially colonize and inhabit a focal habitat area. The concept lays emphasis on the fact that "local communities aren't closed systems, and that the species occupying any local site typically came from somewhere else", however, the species pool concept may suffer from the logical fallacy of composition. Most local communities, however, have just a fraction of its species pool present. It is derived from MacArthur and Wilson's Island Biogeography Theory that examines the factors that affect the species richness of isolated natural communities. It helps to understand the composition and richness of local communities and how they are influenced by biogeographic and evolutionary processes acting at large spatial and temporal scales. The absent portion of species pool—dark diversity—has been used to understand processes influencing local communities. Methods to estimate potential but absent species are developing.