The photic zone, euphotic zone, epipelagic zone, or sunlight zone is the uppermost layer of a body of water that receives sunlight, allowing phytoplankton to perform photosynthesis. It undergoes a series of physical, chemical, and biological processes that supply nutrients into the upper water column. The photic zone is home to the majority of aquatic life due to the activity of the phytoplankton. The thicknesses of the photic and euphotic zones vary with the intensity of sunlight as a function of season and latitude and with the degree of water turbidity. The bottommost, or aphotic, zone is the region of perpetual darkness that lies beneath the photic zone and includes most of the ocean waters.
Phytoplankton are the autotrophic (self-feeding) components of the plankton community and a key part of ocean and freshwater ecosystems. The name comes from the Greek words φυτόν, meaning 'plant', and, meaning 'wanderer' or 'drifter'.
Zooplankton are the animal component of the planktonic community, having to consume other organisms to thrive. Plankton are aquatic organisms that are unable to swim effectively against currents. Consequently, they drift or are carried along by currents in the ocean, or by currents in seas, lakes or rivers.
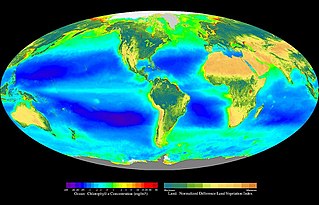
In ecology, primary production is the synthesis of organic compounds from atmospheric or aqueous carbon dioxide. It principally occurs through the process of photosynthesis, which uses light as its source of energy, but it also occurs through chemosynthesis, which uses the oxidation or reduction of inorganic chemical compounds as its source of energy. Almost all life on Earth relies directly or indirectly on primary production. The organisms responsible for primary production are known as primary producers or autotrophs, and form the base of the food chain. In terrestrial ecoregions, these are mainly plants, while in aquatic ecoregions algae predominate in this role. Ecologists distinguish primary production as either net or gross, the former accounting for losses to processes such as cellular respiration, the latter not.
The oxygen minimum zone (OMZ), sometimes referred to as the shadow zone, is the zone in which oxygen saturation in seawater in the ocean is at its lowest. This zone occurs at depths of about 200 to 1,500 m (700–4,900 ft), depending on local circumstances. OMZs are found worldwide, typically along the western coast of continents, in areas where an interplay of physical and biological processes concurrently lower the oxygen concentration and restrict the water from mixing with surrounding waters, creating a "pool" of water where oxygen concentrations fall from the normal range of 4–6 mg/L to below 2 mg/L.

The spring bloom is a strong increase in phytoplankton abundance that typically occurs in the early spring and lasts until late spring or early summer. This seasonal event is characteristic of temperate North Atlantic, sub-polar, and coastal waters. Phytoplankton blooms occur when growth exceeds losses, however there is no universally accepted definition of the magnitude of change or the threshold of abundance that constitutes a bloom. The magnitude, spatial extent and duration of a bloom depends on a variety of abiotic and biotic factors. Abiotic factors include light availability, nutrients, temperature, and physical processes that influence light availability, and biotic factors include grazing, viral lysis, and phytoplankton physiology. The factors that lead to bloom initiation are still actively debated.

A pycnocline is the cline or layer where the density gradient is greatest within a body of water. An ocean current is generated by the forces such as breaking waves, temperature and salinity differences, wind, Coriolis effect, and tides caused by the gravitational pull of celestial bodies. In addition, the physical properties in a pycnocline driven by density gradients also affect the flows and vertical profiles in the ocean. These changes can be connected to the transport of heat, salt, and nutrients through the ocean, and the pycnocline diffusion controls upwelling.
High-nutrient, low-chlorophyll (HNLC) regions are regions of the ocean where the abundance of phytoplankton is low and fairly constant despite the availability of macronutrients. Phytoplankton rely on a suite of nutrients for cellular function. Macronutrients are generally available in higher quantities in surface ocean waters, and are the typical components of common garden fertilizers. Micronutrients are generally available in lower quantities and include trace metals. Macronutrients are typically available in millimolar concentrations, while micronutrients are generally available in micro- to nanomolar concentrations. In general, nitrogen tends to be a limiting ocean nutrient, but in HNLC regions it is never significantly depleted. Instead, these regions tend to be limited by low concentrations of metabolizable iron. Iron is a critical phytoplankton micronutrient necessary for enzyme catalysis and electron transport.
The light compensation point (Ic) is the light intensity on the light curve where the rate of photosynthesis exactly matches the rate of cellular respiration. At this point, the uptake of CO2 through photosynthetic pathways is equal to the respiratory release of carbon dioxide, and the uptake of O2 by respiration is equal to the photosynthetic release of oxygen. The concept of compensation points in general may be applied to other photosynthetic variables, the most important being that of CO2 concentration – CO2 compensation point (Γ).Interval of time in day time when light intensity is low due to which net gaseous exchange is zero is called as compensation point.
Ocean stratification is the natural separation of an ocean's water into horizontal layers by density, which is generally stable because warm water floats on top of cold water, and heating is mostly from the sun, which reinforces that arrangement. Stratification is reduced by wind-forced mechanical mixing, but reinforced by convection. Stratification occurs in all ocean basins and also in other water bodies. Stratified layers are a barrier to the mixing of water, which impacts the exchange of heat, carbon, oxygen and other nutrients. The surface mixed layer is the uppermost layer in the ocean and is well mixed by mechanical (wind) and thermal (convection) effects. Climate change is causing the upper ocean stratification to increase.

Thin layers are concentrated aggregations of phytoplankton and zooplankton in coastal and offshore waters that are vertically compressed to thicknesses ranging from several centimeters up to a few meters and are horizontally extensive, sometimes for kilometers. Generally, thin layers have three basic criteria: 1) they must be horizontally and temporally persistent; 2) they must not exceed a critical threshold of vertical thickness; and 3) they must exceed a critical threshold of maximum concentration. The precise values for critical thresholds of thin layers has been debated for a long time due to the vast diversity of plankton, instrumentation, and environmental conditions. Thin layers have distinct biological, chemical, optical, and acoustical signatures which are difficult to measure with traditional sampling techniques such as nets and bottles. However, there has been a surge in studies of thin layers within the past two decades due to major advances in technology and instrumentation. Phytoplankton are often measured by optical instruments that can detect fluorescence such as LIDAR, and zooplankton are often measured by acoustic instruments that can detect acoustic backscattering such as ABS. These extraordinary concentrations of plankton have important implications for many aspects of marine ecology, as well as for ocean optics and acoustics. Zooplankton thin layers are often found slightly under phytoplankton layers because many feed on them. Thin layers occur in a wide variety of ocean environments, including estuaries, coastal shelves, fjords, bays, and the open ocean, and they are often associated with some form of vertical structure in the water column, such as pycnoclines, and in zones of reduced flow.
The deep chlorophyll maximum (DCM), also called the subsurface chlorophyll maximum, is the region below the surface of water with the maximum concentration of chlorophyll. The DCM generally exists at the same depth as the nutricline, the region of the ocean where the greatest change in the nutrient concentration occurs with depth.

In the deep ocean, marine snow is a continuous shower of mostly organic detritus falling from the upper layers of the water column. It is a significant means of exporting energy from the light-rich photic zone to the aphotic zone below, which is referred to as the biological pump. Export production is the amount of organic matter produced in the ocean by primary production that is not recycled (remineralised) before it sinks into the aphotic zone. Because of the role of export production in the ocean's biological pump, it is typically measured in units of carbon. The term was coined by explorer William Beebe as observed from his bathysphere. As the origin of marine snow lies in activities within the productive photic zone, the prevalence of marine snow changes with seasonal fluctuations in photosynthetic activity and ocean currents. Marine snow can be an important food source for organisms living in the aphotic zone, particularly for organisms that live very deep in the water column.

Hypoxia refers to low oxygen conditions. For air-breathing organisms, hypoxia is problematic but for many anaerobic organisms, hypoxia is essential. Hypoxia applies to many situations, but usually refers to the atmosphere and natural waters.

The lipid pump sequesters carbon from the ocean's surface to deeper waters via lipids associated with overwintering vertically migratory zooplankton. Lipids are a class of hydrocarbon rich, nitrogen and phosphorus deficient compounds essential for cellular structures. This lipid carbon enters the deep ocean as carbon dioxide produced by respiration of lipid reserves and as organic matter from the mortality of zooplankton.

The viral shunt is a mechanism that prevents marine microbial particulate organic matter (POM) from migrating up trophic levels by recycling them into dissolved organic matter (DOM), which can be readily taken up by microorganisms. The DOM recycled by the viral shunt pathway is comparable to the amount generated by the other main sources of marine DOM.

The North Atlantic Aerosols and Marine Ecosystems Study (NAAMES) was a five-year scientific research program that investigated aspects of phytoplankton dynamics in ocean ecosystems, and how such dynamics influence atmospheric aerosols, clouds, and climate. The study focused on the sub-arctic region of the North Atlantic Ocean, which is the site of one of Earth's largest recurring phytoplankton blooms. The long history of research in this location, as well as relative ease of accessibility, made the North Atlantic an ideal location to test prevailing scientific hypotheses in an effort to better understand the role of phytoplankton aerosol emissions on Earth's energy budget.
Shelf-Break Fronts are a process by which stratification of the water column occurs. This stratification normally results in thermoclines, since they occur where a sudden change in water depth causes a constriction of the current flow. They can be expressed as a ratio of their potential energy due to maintaining mixed (non-stratified) conditions, to the dissipated energy produced by the current being forced across the sudden change in depth. This can be expressed as:
Low-nutrient, low-chlorophyll (LNLC)regions are aquatic zones that are low in nutrients and consequently have low rate of primary production, as indicated by low chlorophyll concentrations. These regions can be described as oligotrophic, and about 75% of the world's oceans encompass LNLC regions. A majority of LNLC regions are associated with subtropical gyres but are also present in areas of the Mediterranean Sea, and some inland lakes. Physical processes limit nutrient availability in LNLC regions, which favors nutrient recycling in the photic zone and selects for smaller phytoplankton species. LNLC regions are generally not found near coasts, owing to the fact that coastal areas receive more nutrients from terrestrial sources and upwelling. In marine systems, seasonal and decadal variability of primary productivity in LNLC regions is driven in part by large-scale climatic regimes leading to important effects on the global carbon cycle and the oceanic carbon cycle.
Eddy pumping is a component of mesoscale eddy-induced vertical motion in the ocean. It is a physical mechanism through which vertical motion is created from variations in an eddy's rotational strength. Cyclonic (Anticyclonic) eddies lead primarily to upwelling (downwelling) in the Northern Hemisphere and vice versa in the Southern hemisphere. It is a key mechanism driving biological and biogeochemical processes in the ocean such as algal blooms and the carbon cycle.
