Related Research Articles
The purple sulfur bacteria (PSB) are part of a group of Pseudomonadota capable of photosynthesis, collectively referred to as purple bacteria. They are anaerobic or microaerophilic, and are often found in stratified water environments including hot springs, stagnant water bodies, as well as microbial mats in intertidal zones. Unlike plants, algae, and cyanobacteria, purple sulfur bacteria do not use water as their reducing agent, and therefore do not produce oxygen. Instead, they can use sulfur in the form of sulfide, or thiosulfate (as well, some species can use H2, Fe2+, or NO2−) as the electron donor in their photosynthetic pathways. The sulfur is oxidized to produce granules of elemental sulfur. This, in turn, may be oxidized to form sulfuric acid.

Purple bacteria or purple photosynthetic bacteria are Gram-negative proteobacteria that are phototrophic, capable of producing their own food via photosynthesis. They are pigmented with bacteriochlorophyll a or b, together with various carotenoids, which give them colours ranging between purple, red, brown, and orange. They may be divided into two groups – purple sulfur bacteria and purple non-sulfur bacteria. Purple bacteria are anoxygenic phototrophs widely spread in nature, but especially in aquatic environments, where there are anoxic conditions that favor the synthesis of their pigments.

The Chromatiaceae are one of the two families of purple sulfur bacteria, together with the Ectothiorhodospiraceae. They belong to the order Chromatiales of the class Gammaproteobacteria, which is composed by unicellular Gram-negative organisms. Most of the species are photolithoautotrophs and conduct an anoxygenic photosynthesis, but there are also representatives capable of growing under dark and/or microaerobic conditions as either chemolithoautotrophs or chemoorganoheterotrophs.

Algal mats are one of many types of microbial mat that forms on the surface of water or rocks. They are typically composed of blue-green cyanobacteria and sediments. Formation occurs when alternating layers of blue-green bacteria and sediments are deposited or grow in place, creating dark-laminated layers. Stromatolites are prime examples of algal mats. Algal mats played an important role in the Great Oxidation Event on Earth some 2.3 billion years ago. Algal mats can become a significant ecological problem, if the mats grow so expansive or thick as to disrupt the other underwater marine life by blocking the sunlight or producing toxic chemicals.

Sulfur-reducing bacteria are microorganisms able to reduce elemental sulfur (S0) to hydrogen sulfide (H2S). These microbes use inorganic sulfur compounds as electron acceptors to sustain several activities such as respiration, conserving energy and growth, in absence of oxygen. The final product of these processes, sulfide, has a considerable influence on the chemistry of the environment and, in addition, is used as electron donor for a large variety of microbial metabolisms. Several types of bacteria and many non-methanogenic archaea can reduce sulfur. Microbial sulfur reduction was already shown in early studies, which highlighted the first proof of S0 reduction in a vibrioid bacterium from mud, with sulfur as electron acceptor and H
2 as electron donor. The first pure cultured species of sulfur-reducing bacteria, Desulfuromonas acetoxidans, was discovered in 1976 and described by Pfennig Norbert and Biebel Hanno as an anaerobic sulfur-reducing and acetate-oxidizing bacterium, not able to reduce sulfate. Only few taxa are true sulfur-reducing bacteria, using sulfur reduction as the only or main catabolic reaction. Normally, they couple this reaction with the oxidation of acetate, succinate or other organic compounds. In general, sulfate-reducing bacteria are able to use both sulfate and elemental sulfur as electron acceptors. Thanks to its abundancy and thermodynamic stability, sulfate is the most studied electron acceptor for anaerobic respiration that involves sulfur compounds. Elemental sulfur, however, is very abundant and important, especially in deep-sea hydrothermal vents, hot springs and other extreme environments, making its isolation more difficult. Some bacteria – such as Proteus, Campylobacter, Pseudomonas and Salmonella – have the ability to reduce sulfur, but can also use oxygen and other terminal electron acceptors.

Beggiatoa is a genus of Gammaproteobacteria belonging to the order Thiotrichales, in the Pseudomonadota phylum. These bacteria form colorless filaments composed of cells that can be up to 200 µm in diameter, and are one of the largest prokaryotes on Earth. Beggiatoa are chemolithotrophic sulfur-oxidizers, using reduced sulfur species as an energy source. They live in sulfur-rich environments such as soil, both marine and freshwater, in the deep sea hydrothermal vents, and in polluted marine environments. In association with other sulfur bacteria, e.g. Thiothrix, they can form biofilms that are visible to the naked eye as mats of long white filaments; the white color is due to sulfur globules stored inside the cells.
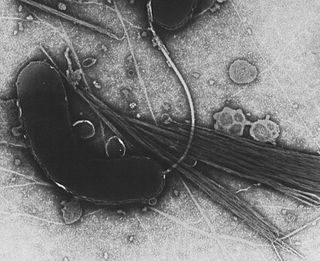
Gammaproteobacteria is a class of bacteria in the phylum Pseudomonadota. It contains about 250 genera, which makes it the most genus-rich taxon of the Prokaryotes. Several medically, ecologically, and scientifically important groups of bacteria belong to this class. All members of this class are Gram-negative. It is the most phylogenetically and physiologically diverse class of the Pseudomonadota.

Phototrophic biofilms are microbial communities generally comprising both phototrophic microorganisms, which use light as their energy source, and chemoheterotrophs. Thick laminated multilayered phototrophic biofilms are usually referred to as microbial mats or phototrophic mats. These organisms, which can be prokaryotic or eukaryotic organisms like bacteria, cyanobacteria, fungi, and microalgae, make up diverse microbial communities that are affixed in a mucous matrix, or film. These biofilms occur on contact surfaces in a range of terrestrial and aquatic environments. The formation of biofilms is a complex process and is dependent upon the availability of light as well as the relationships between the microorganisms. Biofilms serve a variety of roles in aquatic, terrestrial, and extreme environments; these roles include functions which are both beneficial and detrimental to the environment. In addition to these natural roles, phototrophic biofilms have also been adapted for applications such as crop production and protection, bioremediation, and wastewater treatment.

A microbial mat is a multi-layered sheet of microorganisms, mainly bacteria and archaea, or bacteria alone. Microbial mats grow at interfaces between different types of material, mostly on submerged or moist surfaces, but a few survive in deserts. A few are found as endosymbionts of animals.
The Sippewissett microbial mat is a microbial mat in the Sippewissett Salt Marsh located along the lower eastern Buzzards Bay shoreline of Cape Cod, about 5 miles north of Woods Hole and 1 mile southwest of West Falmouth, Massachusetts, in the United States. The marsh has two regions, the Great Sippewisset Marsh to the north and Little Sippewisset Marsh to the south, separated from each other by a narrow tongue of land. The marsh extends into an estuary in which the intertidal zone provides a dynamic environment that supports a diverse ecology, including threatened and endangered species such as the roseate tern. The ecology of the salt marsh is based in and supported by the microbial mats which cover the ground of the marsh.
Rhodovulum sulfidophilum is a gram-negative purple nonsulfur bacteria. The cells are rod-shaped, and range in size from 0.6 to 0.9 μm wide and 0.9 to 2.0 μm long, and have a polar flagella. These cells reproduce asexually by binary fission. This bacterium can grow anaerobically when light is present, or aerobically (chemoheterotrophic) under dark conditions. It contains the photosynthetic pigments bacteriochlorophyll a and of carotenoids.
Roseiflexus castenholzii is a heterotrophic, thermophilic, filamentous anoxygenetic phototroph (FAP) bacterium. This species is in one of two genera of FAPs that lack chlorosomes. R. castenholzii was first isolated from red-colored bacterial mats located Nakabusa hot springs in Japan. Because this organism is a phototroph, it utilizes photosynthesis to fix carbon dioxide and build biomolecules. R. castenholzii has three photosynthetic complexes: light-harvesting only, reaction center only, and light-harvesting with reaction center.
Rhodoferax is a genus of Betaproteobacteria belonging to the purple nonsulfur bacteria. Originally, Rhodoferax species were included in the genus Rhodocyclus as the Rhodocyclus gelatinous-like group. The genus Rhodoferax was first proposed in 1991 to accommodate the taxonomic and phylogenetic discrepancies arising from its inclusion in the genus Rhodocyclus. Rhodoferax currently comprises four described species: R. fermentans, R. antarcticus, R. ferrireducens, and R. saidenbachensis. R. ferrireducens, lacks the typical phototrophic character common to two other Rhodoferax species. This difference has led researchers to propose the creation of a new genus, Albidoferax, to accommodate this divergent species. The genus name was later corrected to Albidiferax. Based on geno- and phenotypical characteristics, A. ferrireducens was reclassified in the genus Rhodoferax in 2014. R. saidenbachensis, a second non-phototrophic species of the genus Rhodoferax was described by Kaden et al. in 2014.
Thiorhodovibrio winogradskyi is a purple sulfur bacteria, the type species of its genus. Its cells are vibrioid-to spirilloid-shaped and motile by means of single polar flagella. It is moderately halophilic, with type strain SSP1.
Halorhodospira halophila is a species of Halorhodospira distinguished by its ability to grow optimally in an environment of 15–20% salinity. It was formerly called Ectothiorhodospira halophila. It is an anaerobic, rod-shaped Gram-negative bacterium. H. halophila has a flagellum.
Dinoroseobacter shibae is a facultative anaerobic anoxygenic photoheterotroph belonging to the family, Rhodobacteraceae. First isolated from washed cultivated dinoflagellates, they have been reported to have mutualistic as well as pathogenic symbioses with dinoflagellates.
Thioflavicoccus is a Gram-negative, obligately phototrophic, strictly anaerobic and motile genus of bacteria from the family of Chromatiaceae with one known species.
Rhodoplanes azumiensis is a thermotolerant bacterium isolated from sediment mud and cyanobacterial mats located in Nakanoyu hot springs, located in the Azumi district, Matsumoto, Nagano Prefecture, Japan. R. azumiensis is a photoheterotroph that is capable of using many organic materials as its carbon source. Colonies and liquid cultures exhibit a brownish red color when grown phototrophically, and are colorless when grown aerobically in darkness.
Thiodictyon is a genus of gram-negative bacterium classified within purple sulfur bacteria (PSB).
Jeotgalibaca is a genus of bacteria from the family of Carnobacteriaceae.
References
- 1 2 "Halorhodospira". LPSN .
- 1 2 3 "Halorhodospira neutriphila". www.uniprot.org.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 Hirschler-Réa, A; Matheron, R; Riffaud, C; Mouné, S; Eatock, C; Herbert, RA; Willison, JC; Caumette, P (January 2003). "Isolation and characterization of spirilloid purple phototrophic bacteria forming red layers in microbial mats of Mediterranean salterns: description of Halorhodospira neutriphila sp. nov. and emendation of the genus Halorhodospira". International Journal of Systematic and Evolutionary Microbiology. 53 (Pt 1): 153–63. doi: 10.1099/ijs.0.02226-0 . PMID 12656167.
- ↑ Parker, Charles Thomas; Garrity, George M (19 January 2017). Parker, Charles Thomas; Garrity, George M (eds.). "Taxonomic Abstract for the species". NamesforLife, LLC. doi:10.1601/tx.2175 (inactive 2024-04-17).
{{cite journal}}: Cite journal requires|journal=(help)CS1 maint: DOI inactive as of April 2024 (link) - ↑ "Details: DSM-15116". www.dsmz.de.
- ↑ Sayers; et al. (2018). "National Center for Biotechnological Information". NCBI.