
Dentition pertains to the development of teeth and their arrangement in the mouth. In particular, it is the characteristic arrangement, kind, and number of teeth in a given species at a given age. That is, the number, type, and morpho-physiology of the teeth of an animal.

In mammalian oral anatomy, the canine teeth, also called cuspids, dog teeth, or fangs, eye teeth, vampire teeth, or vampire fangs, are the relatively long, pointed teeth. They can appear more flattened however, causing them to resemble incisors and leading them to be called incisiform. They developed and are used primarily for firmly holding food in order to tear it apart, and occasionally as weapons. They are often the largest teeth in a mammal's mouth. Individuals of most species that develop them normally have four, two in the upper jaw and two in the lower, separated within each jaw by incisors; humans and dogs are examples. In most species, canines are the anterior-most teeth in the maxillary bone.

Catopsbaatar is a genus of multituberculate, an extinct order of rodent-like mammals. It lived in what is now Mongolia during the late Campanian age of the Late Cretaceous epoch, about 72 million years ago. The first fossils were collected in the early 1970s, and the animal was named as a new species of the genus Djadochtatherium in 1974, D. catopsaloides. The specific name refers to the animal's similarity to the genus Catopsalis. The species was moved to the genus Catopsalis in 1979, and received its own genus in 1994. Five skulls, one molar, and one skeleton with a skull are known; the last is the genus' most complete specimen. Catopsbaatar was a member of the family Djadochtatheriidae.
Ferugliotherium is a genus of fossil mammals in the family Ferugliotheriidae from the Campanian and/or Maastrichtian period of Argentina. It contains a single species, Ferugliotherium windhauseni, which was first described in 1986. Although originally interpreted on the basis of a single brachydont (low-crowned) molar as a member of Multituberculata, an extinct group of small, rodent-like mammals, it was recognized as related to the hypsodont (high-crowned) Sudamericidae following the discovery of additional material in the early 1990s. After a jaw of the sudamericid Sudamerica was described in 1999, these animals were no longer considered to be multituberculates and a few fossils that were previously considered to be Ferugliotherium were assigned to unspecified multituberculates instead. Since 2005, a relationship between gondwanatheres and multituberculates has again received support. A closely related animal, Trapalcotherium, was described in 2009 on the basis of a single tooth.

Incisors are the front teeth present in most mammals. They are located in the premaxilla above and on the mandible below. Humans have a total of eight. Opossums have 18, whereas armadillos have none.

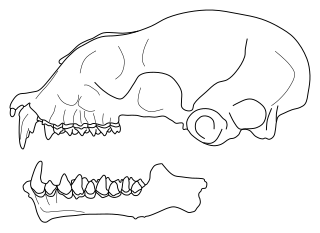
Gigantopithecus is an extinct genus of ape from the Early to Middle Pleistocene of southern China, represented by one species, G. blacki. Potential identifications have also been made in Thailand, Vietnam, and Indonesia. The first remains of Gigantopithecus, two third molar teeth, were identified in a drugstore by anthropologist Ralph von Koenigswald in 1935, who subsequently described the ape. In 1956, the first mandible and over 1,000 teeth were found in Liucheng, and numerous more remains have since been found in at least 16 sites. Only teeth and four mandibles are known currently, and other skeletal elements were likely consumed by porcupines before they could fossilise. Gigantopithecus was once argued to be a hominin, a member of the human line, but it is now thought to be closely allied with orangutans, classified in the subfamily Ponginae.
Ferugliotheriidae is one of three known families in the order Gondwanatheria, an enigmatic group of extinct mammals. Gondwanatheres have been classified as a group of uncertain affinities or as members of Multituberculata, a major extinct mammalian order. The best-known representative of Ferugliotheriidae is the genus Ferugliotherium from the Late Cretaceous epoch in Argentina. A second genus, Trapalcotherium, is known from a single tooth, a first lower molariform, from a different Late Cretaceous Argentinean locality. Another genus known from a single tooth, Argentodites, was first described as an unrelated multituberculate, but later identified as possibly related to Ferugliotherium. Finally, a single tooth from the Paleogene of Peru, LACM 149371, perhaps a last upper molariform, and a recent specimen from Mexico, may represent related animals.

The dental arches are the two arches of teeth, one on each jaw, that together constitute the dentition. In humans and many other species; the superior dental arch is a little larger than the inferior arch, so that in the normal condition the teeth in the maxilla slightly overlap those of the mandible both in front and at the sides. The way that the jaws, and thus the dental arches, approach each other when the mouth closes, which is called the occlusion, determines the occlusal relationship of opposing teeth, and it is subject to malocclusion if facial or dental development was imperfect.
Dental anatomy is a field of anatomy dedicated to the study of human tooth structures. The development, appearance, and classification of teeth fall within its purview. Tooth formation begins before birth, and the teeth's eventual morphology is dictated during this time. Dental anatomy is also a taxonomical science: it is concerned with the naming of teeth and the structures of which they are made, this information serving a practical purpose in dental treatment.

True's shrew mole is a species of mammal in the family Talpidae. It is endemic to Japan and is a common species above 1000 meters in grassland, shrubland and forest.

Eritherium is an extinct genus of early Proboscidea found in the Ouled Abdoun basin, Morocco. It lived about 60 million years ago. It was first named by Emmanuel Gheerbrant in 2009 and the type species is Eritherium azzouzorum. Eritherium is the oldest, smallest and most primitive known elephant relative.

Microgale macpheei is an extinct shrew tenrec from southeastern Madagascar. It is known only from two partial skulls found in Andrahomana cave, which radiocarbon dating of associated rodent remains suggests are about 3000 years old. It is the only known recently extinct tenrec. First described in 2007, it is most similar to the smaller Microgale brevicaudata of northern and western Madagascar. M. macpheei has a broad rostrum and, like M. brevicaudata, lacks a diastema (gap) between the premolars. A number of details of tooth morphology are characteristic of M. macpheei.

Triaenops goodmani is an extinct bat from Madagascar in the genus Triaenops. It is known from three lower jaws collected in a cave at Anjohibe in 1996, and described as a new species in 2007. The material is at most 10,000 years old. A bat humerus from the same site could not be identified as either T. goodmani or the living T. menamena. T. goodmani is identifiable as a member of Triaenops or the related genus Paratriaenops by a number of features of the teeth, such as the single-cusped, canine-like fourth premolar and the presence of a gap between the entoconid and hypoconulid cusps on the first two molars. T. goodmani is larger than the living species of Triaenops and Paratriaenops on Madagascar, and on the first molar the protoconid cusp is only slightly higher than the hypoconid, not much higher as in the other species.
Triaenops menamena is a bat in the genus Triaenops found on Madagascar, mainly in the drier regions. It was known as Triaenops rufus until 2009, when it was discovered that that name had been incorrectly applied to the species. Triaenops rufus is a synonym of Triaenops persicus, a Middle Eastern species closely related to T. menamena— the Malagasy species had previously been placed as a subspecies of T. persicus by some authors. Triaenops menamena is mostly found in forests, but also occurs in other habitats. It often roosts in large colonies and eats insects such as butterflies and moths. Because of its wide range, common occurrence, and tolerance of habitat degradation, it is not considered to be threatened.

Pipistrellus raceyi, also known as Racey's pipistrelle, is a bat from Madagascar, in the genus Pipistrellus. Although unidentified species of Pipistrellus had been previously reported from Madagascar since the 1990s, P. raceyi was not formally named until 2006. It is apparently most closely related to the Asian species P. endoi, P. paterculus, and P. abramus, and its ancestors probably reached Madagascar from Asia. P. raceyi has been recorded at four sites, two in the eastern and two in the western lowlands. In the east, it is found in open areas and has been found roosting in a building; in the west it occurs in dry forest. Because of uncertainties about its ecology, it is listed as "Data Deficient" on the IUCN Red List.

Durrell's vontsira is a Madagascan mammal in the family Eupleridae of the order Carnivora. It is most closely related to the brown-tailed mongoose, with which it forms the genus Salanoia. The two are genetically similar, but morphologically distinct, leading scientists to recognize them as separate species. After an individual was observed in 2004, the animal became known to science and S. durrelli was described as a new species in 2010. It is found only in the Lac Alaotra area.

Teeth are common to most vertebrates, but mammalian teeth are distinctive in having a variety of shapes and functions. This feature first arose among early therapsids during the Permian, and has continued to the present day. All therapsid groups with the exception of the mammals are now extinct, but each of these groups possessed different tooth patterns, which aids with the classification of fossils.
Dermotherium is a genus of fossil mammals closely related to the living colugos, a small group of gliding mammals from Southeast Asia. Two species are recognized: D. major from the Late Eocene of Thailand, based on a single fragment of the lower jaw, and D. chimaera from the Late Oligocene of Thailand, known from three fragments of the lower jaw and two isolated upper molars. In addition, a single isolated upper molar from the Early Oligocene of Pakistan has been tentatively assigned to D. chimaera. All sites where fossils of Dermotherium have been found were probably forested environments and the fossil species were probably forest dwellers like living colugos, but whether they had the gliding adaptations of the living species is unknown.
Changes to the dental morphology and jaw are major elements of hominid evolution. These changes were driven by the types and processing of food eaten. The evolution of the jaw is thought to have facilitated encephalization, speech, and the formation of the uniquely human chin.
Dentition analyses are systems of tooth and jaw measurement used in orthodontics to understand arch space and predict any malocclusion. Example systems of dentition analysis are listed below.
