
A herbivore is an animal anatomically and physiologically adapted to eating plant material, for example foliage or marine algae, for the main component of its diet. As a result of their plant diet, herbivorous animals typically have mouthparts adapted to rasping or grinding. Horses and other herbivores have wide flat teeth that are adapted to grinding grass, tree bark, and other tough plant material.
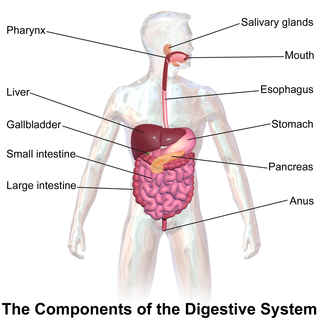
Digestion is the breakdown of large insoluble food molecules into small water-soluble food molecules so that they can be absorbed into the watery blood plasma. In certain organisms, these smaller substances are absorbed through the small intestine into the blood stream. Digestion is a form of catabolism that is often divided into two processes based on how food is broken down: mechanical and chemical digestion. The term mechanical digestion refers to the physical breakdown of large pieces of food into smaller pieces which can subsequently be accessed by digestive enzymes. Mechanical digestion takes place in the mouth through mastication and in the small intestine through segmentation contractions. In chemical digestion, enzymes break down food into the small molecules the body can use.

The cedar waxwing is a member of the family Bombycillidae or waxwing family of passerine birds. It is a medium-sized, mostly brown, gray, and yellow. This bird is named for its wax-like wing tips. It is a native of North and Central America, breeding in open wooded areas in southern Canada and wintering in the southern half of the United States, Central America, and the far northwest of South America. Its diet includes cedar cones, fruit, holly berries, and insects. The cedar waxwing is listed as least concern on the IUCN Red List.

Eating is the ingestion of food, typically to provide a heterotrophic organism with energy and to allow for growth. Animals and other heterotrophs must eat in order to survive — carnivores eat other animals, herbivores eat plants, omnivores consume a mixture of both plant and animal matter, and detritivores eat detritus. Fungi digest organic matter outside their bodies as opposed to animals that digest their food inside their bodies. For humans, eating is an activity of daily living. Some individuals may limit their amount of nutritional intake. This may be a result of a lifestyle choice, due to hunger or famine, as part of a diet or as religious fasting.

Ruminants are hoofed herbivorous grazing or browsing mammals that are able to acquire nutrients from plant-based food by fermenting it in a specialized stomach prior to digestion, principally through microbial actions. The process, which takes place in the front part of the digestive system and therefore is called foregut fermentation, typically requires the fermented ingesta to be regurgitated and chewed again. The process of rechewing the cud to further break down plant matter and stimulate digestion is called rumination. The word "ruminant" comes from the Latin ruminare, which means "to chew over again".

Chewing or mastication is the process by which food is crushed and ground by teeth. It is the first step of digestion, and it increases the surface area of foods to allow a more efficient break down by enzymes. During the mastication process, the food is positioned by the cheek and tongue between the teeth for grinding. The muscles of mastication move the jaws to bring the teeth into intermittent contact, repeatedly occluding and opening. As chewing continues, the food is made softer and warmer, and the enzymes in saliva begin to break down carbohydrates in the food. After chewing, the food is swallowed. It enters the esophagus and via peristalsis continues on to the stomach, where the next step of digestion occurs. Increasing the number of chews per bite increases relevant gut hormones. Studies suggest that chewing may decrease self-reported hunger and food intake. Chewing gum has been around for many centuries; there is evidence that northern Europeans chewed birch bark tar 9,000 years ago.
A monogastric organism has a simple single-chambered stomach. Examples of monogastric herbivores are horses and rabbits. Examples of monogastric omnivores include humans, pigs, hamsters and rats. Furthermore, there are monogastric carnivores such as cats. A monogastric organism is comparable to ruminant organisms, such as cattle, goats, or sheep. Herbivores with monogastric digestion can digest cellulose in their diets by way of symbiotic gut bacteria. However, their ability to extract energy from cellulose digestion is less efficient than in ruminants.

In zoology, a folivore is a herbivore that specializes in eating leaves. Mature leaves contain a high proportion of hard-to-digest cellulose, less energy than other types of foods, and often toxic compounds. For this reason, folivorous animals tend to have long digestive tracts and slow metabolisms. Many enlist the help of symbiotic bacteria to release the nutrients in their diet. Additionally, as has been observed in folivorous primates, they exhibit a strong preference for immature leaves, which tend to be easier to masticate, tend to be higher in energy and protein, and lower in fibre and poisons than more mature fibrous leaves.
The rumen, also known as a paunch, is the largest stomach compartment in ruminants and the larger part of the reticulorumen, which is the first chamber in the alimentary canal of ruminant animals. The rumen's microbial favoring environment allows it to serve as the primary site for microbial fermentation of ingested feed. The smaller part of the reticulorumen is the reticulum, which is fully continuous with the rumen, but differs from it with regard to the texture of its lining.

Macronaria is a clade of sauropod dinosaurs. Macronarians are named after the large diameter of the nasal opening of their skull, known as the external naris, which exceeded the size of the orbit, the skull opening where the eye is located. Fossil evidence suggests that macronarian dinosaurs lived from the Middle Jurassic (Bathonian) through the Late Cretaceous (Maastrichtian). Macronarians have been found globally, including discoveries in Argentina, the United States, Portugal, China, and Tanzania. Like other sauropods, they are known to have inhabited primarily terrestrial areas, and little evidence exists to suggest that they spent much time in coastal environments. Macronarians are diagnosed through their distinct characters on their skulls, as well as appendicular and vertebral characters. Macronaria is composed of several subclades and families notably including Camarasauridae and Titanosauriformes, among several others. Titanosauriforms are particularly well known for being some of the largest terrestrial animals to ever exist.

Size is an important aspect of dinosaur paleontology, of interest to both the general public and professional scientists. Dinosaurs show some of the most extreme variations in size of any land animal group, ranging from tiny hummingbirds, which can weigh as little as two grams, to the extinct titanosaurs, which could weigh as much as 50–100 t.
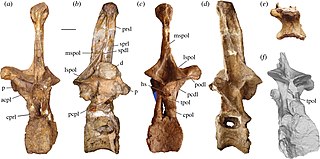
Amphicoelias is a genus of herbivorous sauropod dinosaur that lived approximately 150 million years ago during the Tithonian of what is now Colorado, United States. Amphicoelias was moderately sized at about 18 metres (59 ft) in length and 15 metric tons in body mass, shorter than its close relative Diplodocus. Its hindlimbs were very long and thin, and its forelimbs were proportionally longer than in relatives.
The digestive rate model (DRM) is related to optimal foraging theory in that the model describes the diet selection that animals should perform in order to maximize the energy available to them. It differs from the main body of Optimal Foraging Theory in stating that animals can select food in order to make optimal use of their digestive tract rather than the maximization of the food ingestion rate, which is the base of Optimal foraging theory.
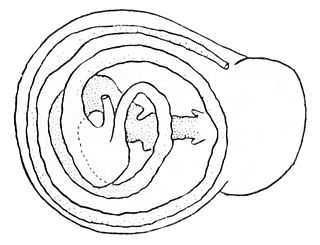
The digestive system of gastropods has evolved to suit almost every kind of diet and feeding behavior. Gastropods as the largest taxonomic class of the mollusca are very diverse: the group includes carnivores, herbivores, scavengers, filter feeders, and even parasites.
Hindgut fermentation is a digestive process seen in monogastric herbivores, animals with a simple, single-chambered stomach. Cellulose is digested with the aid of symbiotic bacteria. The microbial fermentation occurs in the digestive organs that follow the small intestine: the large intestine and cecum. Examples of hindgut fermenters include proboscideans and large odd-toed ungulates such as horses and rhinos, as well as small animals such as rodents, rabbits and koalas. In contrast, foregut fermentation is the form of cellulose digestion seen in ruminants such as cattle which have a four-chambered stomach, as well as in sloths, macropodids, some monkeys, and one bird, the hoatzin.
A gummivore is an omnivorous animal whose diet consists primarily of the gums and saps of trees and bugs for protein. Notable gummivores include arboreal, terrestrial primates like certain marmosets and lemurs. These animals that live off of the injuries of trees live from about 8m off of the ground up to the canopies. The feeding habit of gummivores is gummivory.

Maraapunisaurus is a genus of sauropod dinosaur from the Late Jurassic Morrison Formation of western North America. It is known only from what has sometimes been estimated to be the largest dinosaur specimen ever discovered, originally named Amphicoelias fragillimus. Based on surviving descriptions of a single fossil bone, scientists have produced numerous size estimates over the years; the largest estimate M. fragillimus to have been the longest known animal at 58 metres (190 ft) in length and with a mass of 150 tonnes. However, because the only fossil remains were lost at some point after being studied and described in the 1870s, evidence survived only in contemporary drawings and field notes.
The expensive tissue hypothesis (ETH) relates brain and gut size in evolution. It suggests that in order for an organism to evolve a large brain without a significant increase in basal metabolic rate, the organism must use less energy on other expensive tissues; the paper introducing the ETH suggests that in humans, this was achieved by eating an easy-to-digest diet and evolving a smaller, less energy intensive gut. The ETH has inspired many research projects to test its validity in primates and other organisms.

Anaeromyces robustus is a fungal microorganism that lives in the gut rumen of many ruminant herbivores such as cows and sheep. Previously thought to be protozoa from their flagellated zoospores, they are biomass degraders and help the animal by breaking down carbohydrates and plant materials from the food the animal ingests. This fungus, therefore, is anaerobic and lives without oxygen. Gut fungi are dramatically outnumbered by other organisms in the microbiome; however, they are important members of the gut microbiome in ruminants and hind-gut fermenters and play a key role in digestion.

Megaherbivores are large terrestrial herbivores that can exceed 1,000 kg (2,200 lb) in weight. This polyphyletic group of megafauna includes elephants, rhinos, hippos, and giraffes. The largest bovids occasionally reach a weight of 1 tonne, but they are generally not considered to be megaherbivores. There are nine extant species of megaherbivores living in Africa and Asia. The African bush elephant is the largest extant species with bulls reaching a height of up to 3.96 m (13.0 ft) and a maximum weight of 10,400 kg (22,900 lb).











