Interferons are a group of signaling proteins made and released by host cells in response to the presence of several viruses. In a typical scenario, a virus-infected cell will release interferons causing nearby cells to heighten their anti-viral defenses.

Dengue virus (DENV) is the cause of dengue fever. It is a mosquito-borne, single positive-stranded RNA virus of the family Flaviviridae; genus Flavivirus. Four serotypes of the virus have been found, and a reported fifth has yet to be confirmed, all of which can cause the full spectrum of disease. Nevertheless, the mainstream scientific community's understanding of dengue virus may be simplistic as, rather than distinct antigenic groups, a continuum appears to exist. This same study identified 47 strains of dengue virus. Additionally, coinfection with and lack of rapid tests for Zika virus and chikungunya complicate matters in real-world infections.

The innate immune system or nonspecific immune system is one of the two main immunity strategies in vertebrates. The innate immune system is an alternate defense strategy and is the dominant immune system response found in plants, fungi, prokaryotes, and invertebrates.

Caspase recruitment domains, or caspase activation and recruitment domains (CARDs), are interaction motifs found in a wide array of proteins, typically those involved in processes relating to inflammation and apoptosis. These domains mediate the formation of larger protein complexes via direct interactions between individual CARDs. CARDs are found on a strikingly wide range of proteins, including helicases, kinases, mitochondrial proteins, caspases, and other cytoplasmic factors.

Interferon regulatory factor 3, also known as IRF3, is an interferon regulatory factor.

Toll-like receptor 6 is a protein that in humans is encoded by the TLR6 gene. TLR6 is a transmembrane protein, member of toll-like receptor family, which belongs to the pattern recognition receptor (PRR) family. TLR6 acts in a heterodimer form with toll-like receptor 2 (TLR2). Its ligands include multiple diacyl lipopeptides derived from gram-positive bacteria and mycoplasma and several fungal cell wall saccharides. After dimerizing with TLR2, the NF-κB intracellular signalling pathway is activated, leading to a pro-inflammatory cytokine production and activation of innate immune response. TLR6 has also been designated as CD286.

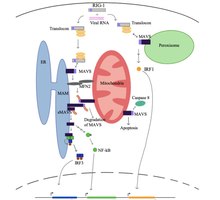
RIG-I is a cytosolic pattern recognition receptor (PRR) that can mediate induction of a type-I interferon (IFN1) response. RIG-I is an essential molecule in the innate immune system for recognizing cells that have been infected with a virus. These viruses can include West Nile virus, Japanese Encephalitis virus, influenza A, Sendai virus, flavivirus, and coronaviruses.

TBK1 is an enzyme with kinase activity. Specifically, it is a serine / threonine protein kinase. It is encoded by the TBK1 gene in humans. This kinase is mainly known for its role in innate immunity antiviral response. However, TBK1 also regulates cell proliferation, apoptosis, autophagy, and anti-tumor immunity. Insufficient regulation of TBK1 activity leads to autoimmune, neurodegenerative diseases or tumorigenesis.

Interferon regulatory factor 5 is a protein that in humans is encoded by the IRF5 gene. The IRF family is a group of transcription factors that are involved in signaling for virus responses in mammals along with regulation of certain cellular functions.

MDA5 is a RIG-I-like receptor dsRNA helicase enzyme that is encoded by the IFIH1 gene in humans. MDA5 is part of the RIG-I-like receptor (RLR) family, which also includes RIG-I and LGP2, and functions as a pattern recognition receptor capable of detecting viruses. It is generally believed that MDA5 recognizes double stranded RNA (dsRNA) over 2000nts in length, however it has been shown that whilst MDA5 can detect and bind to cytoplasmic dsRNA, it is also activated by a high molecular weight RNA complex composed of ssRNA and dsRNA. For many viruses, effective MDA5-mediated antiviral responses are dependent on functionally active LGP2. The signaling cascades in MDA5 is initiated via CARD domain. Some observations made in cancer cells show that MDA5 also interacts with cellular RNA is able to induce an autoinflammatory response.
Mitophagy is the selective degradation of mitochondria by autophagy. It often occurs to defective mitochondria following damage or stress. The process of mitophagy was first described in 1915 by Margaret Reed Lewis and Warren Harmon Lewis. Ashford and Porter used electron microscopy to observe mitochondrial fragments in liver lysosomes by 1962, and a 1977 report suggested that "mitochondria develop functional alterations which would activate autophagy." The term "mitophagy" was in use by 1998.
RIG-I-like receptors are a type of intracellular pattern recognition receptor involved in the recognition of viruses by the innate immune system. RIG-I is the best characterized receptor within the RIG-I like receptor (RLR) family. Together with MDA5 and LGP2, this family of cytoplasmic pattern recognition receptors (PRRs) are sentinels for intracellular viral RNA that is a product of viral infection. The RLR receptors provide frontline defence against viral infections in most tissues.

Stimulator of interferon genes (STING), also known as transmembrane protein 173 (TMEM173) and MPYS/MITA/ERIS is a protein that in humans is encoded by the STING1 gene.
Shu Hongbing is a Chinese cytologist and immunologist. He became a member of the Chinese Academy of Sciences in 2011 and TWAS in 2012. Shu is mainly known for his work on cell signal transduction related to immunity.

Mitochondrial E3 ubiquitin protein ligase 1 (MUL1) is an enzyme that in humans is encoded by the MUL1 gene on chromosome 1. This enzyme localizes to the outer mitochondrial membrane, where it regulates mitochondrial morphology and apoptosis through multiple pathways, including the Akt, JNK, and NF-κB. Its proapoptotic function thus implicates it in cancer and Parkinson's disease.
The cGAS–STING pathway is a component of the innate immune system that functions to detect the presence of cytosolic DNA and, in response, trigger expression of inflammatory genes that can lead to senescence or to the activation of defense mechanisms. DNA is normally found in the nucleus of the cell. Localization of DNA to the cytosol is associated with tumorigenesis, viral infection, and invasion by some intracellular bacteria. The cGAS – STING pathway acts to detect cytosolic DNA and induce an immune response.

Evolutionarily conserved signaling intermediate in Toll pathway, mitochondrial (ECSIT), also known as SITPEC, is a protein that in humans is encoded by the ECSIT gene. ECSIT is a cytosolic adaptor protein involved in inflammatory responses, embryonic development, and the assembly and stabilization of mitochondrial NADH:ubiquinone oxidoreductase.

The SNX8 is a sorting nexin protein involved in intracellular molecular traffic from the early endosomes to the trans-golgi network. It is suggested that it acts as an adaptor protein in events related to immune response and cholesterol regulation, for example. As a protein of the SNXs family, the SNX8 is formed of 465 aminoacids and presents a BAR domain and a PX domain which are very relevant in relation to its functions. Furthermore, SNX8 study is motivated by its medical significance in relation to diseases such as Alzheimer's Disease, cancer, neurodevelopmental malformations and to its role in fighting against viral infections.

The mammalian immune system has evolved complex methods for addressing and adapting to foreign antigens. At the same time, viruses have co-evolved evasion machinery to address the many ways that host organisms attempt to eradicate them. DNA and RNA viruses use complex methods to evade immune cell detection through disruption of the Interferon Signaling Pathway, remodeling of cellular architecture, targeted gene silencing, and recognition protein cleavage.
Jonathan C. Kagan is an American immunologist and the Marian R. Neutra, Ph.D. Professor of Pediatrics at Harvard Medical School. He is also the director of Basic Research and Shwachman Chair in Gastroenterology at Boston Children's Hospital. Kagan is a world leader in defining the molecular basis of innate immunity and inflammation.