
The Bombyliidae are a family of flies, commonly known as bee flies. Some are colloquially known as bomber flies. Adults generally feed on nectar and pollen, some being important pollinators. Larvae are mostly parasitoids of other insects.

The Phoridae are a family of small, hump-backed flies resembling fruit flies. Phorid flies can often be identified by their escape habit of running rapidly across a surface rather than taking flight. This behaviour is a source of one of their alternate names, scuttle fly. Another vernacular name, coffin fly, refers to Conicera tibialis. About 4,000 species are known in 230 genera. The most well-known species is cosmopolitan Megaselia scalaris. At 0.4 mm in length, the world's smallest fly is the phorid Euryplatea nanaknihali.

The Conopidae, also known as the thick-headed flies, are a family of flies within the Brachycera suborder of Diptera, and the sole member of the superfamily Conopoidea. Flies of the family Conopidae are distributed worldwide in all the biogeographic realms except for the poles and many of the Pacific islands. About 800 species in 47 genera are described worldwide, about 70 of which are found in North America. The majority of conopids are black and yellow, or black and white, and often strikingly resemble wasps, bees, or flies of the family Syrphidae, themselves notable bee mimics. A conopid is most frequently found at flowers, feeding on nectar with its proboscis, which is often long.

The Therevidae are a family of flies of the superfamily Asiloidea commonly known as stiletto flies. The family contains about 1,600 described species worldwide, most diverse in arid and semiarid regions with sandy soils. The larvae are predators of insect larvae in soil.

The soldier flies are a family of flies. The family contains over 2,700 species in over 380 extant genera worldwide. Larvae are found in a wide array of locations, mostly in wetlands, damp places in soil, sod, under bark, in animal excrement, and in decaying organic matter. Adults are found near larval habitats. They are diverse in size and shape, though they commonly are partly or wholly metallic green, or somewhat wasplike mimics, marked with black and yellow or green and sometimes metallic. They are often rather inactive flies which typically rest with their wings placed one above the other over the abdomen.

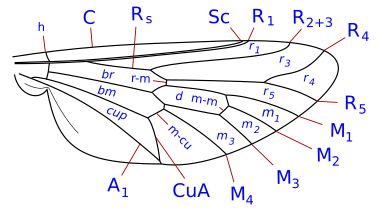
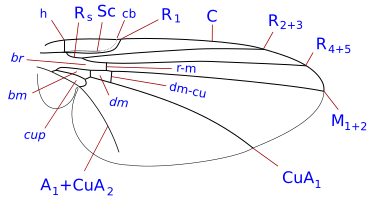
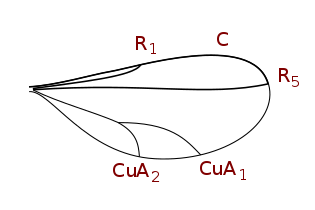
Insect wings are adult outgrowths of the insect exoskeleton that enable insects to fly. They are found on the second and third thoracic segments, and the two pairs are often referred to as the forewings and hindwings, respectively, though a few insects lack hindwings, even rudiments. The wings are strengthened by a number of longitudinal veins, which often have cross-connections that form closed "cells" in the membrane. The patterns resulting from the fusion and cross-connection of the wing veins are often diagnostic for different evolutionary lineages and can be used for identification to the family or even genus level in many orders of insects.

This glossary of entomology describes terms used in the formal study of insect species by entomologists.

The Agromyzidae are a family of flies, commonly referred to as the leaf-miner flies for the feeding habits of their larvae, most of which are leaf miners on various plants. It includes roughly 2,500 species, they are small, some with wing length of 1 mm. The maximum size is 6.5 mm. Most species are in the range of 2 to 3 mm.

Empididae is a family of flies with over 3,000 described species occurring worldwide in all the biogeographic realms but the majority are found in the Holarctic. They are mainly predatory flies like most of their relatives in the Empidoidea, and exhibit a wide range of forms but are generally small to medium-sized, non-metallic and rather bristly.

The Anisopodidae are a small cosmopolitan family of gnat-like flies known as wood gnats or window-gnats, with 154 described extant species in 15 genera, and several described fossil taxa. Some species are saprophagous or fungivorous. They are mostly small to medium-sized flies, except the genera Olbiogaster and Lobogaster, which are large with bizarrely spatulated abdomens. Their phylogenetic placement is controversial. They have been proposed to be the sister group to the higher flies, the Brachycera. Some authors consider this group to be four distinct families – Anisopodidae, Mycetobiidae, Olbiogastridae, and Valeseguyidae.

The Lonchaeidae are a family of acalyptrate flies commonly known as lance flies. 610 described species are placed into 10 genera. These are generally small but robustly built flies with blue-black or metallic bodies. They are found, most commonly in wooded areas, throughout the world with the exception of polar regions and New Zealand. Details of the distribution of genera and species by biogeographic realm are included in the World Catalogue of the family Lonchaeidae

Diastatidae are a family of flies in the order Diptera. They are encountered primarily in the Holarctic Region, but several species are found in the Oriental, Neotropical and Australasian regions. Members of the family number over 20 described species in three genera. There is an additional fossil genus.

Pallopteridae is a family of flies. The various species are collectively called flutter-wing flies, trembling-wing, or waving-wing flies, because of the striking vibration of the wings in many species. Over 70 species in about 15 genera are found in the temperate regions of the Northern and Southern Hemispheres.
The Scenopinidae or window flies are a small family of flies (Diptera), distributed worldwide. In buildings, they are often taken at windows, hence the common name window flies.

The Dixidae are a family of aquatic nematoceran flies (Diptera). The larvae live in unpolluted, standing fresh waters, just beneath the surface film, usually amongst marginal aquatic vegetation. They are found in all continents except Antarctica.

Insect morphology is the study and description of the physical form of insects. The terminology used to describe insects is similar to that used for other arthropods due to their shared evolutionary history. Three physical features separate insects from other arthropods: they have a body divided into three regions, three pairs of legs, and mouthparts located outside of the head capsule. This position of the mouthparts divides them from their closest relatives, the non-insect hexapods, which include Protura, Diplura, and Collembola.
Odonata are insects with an incomplete metamorphosis (hemimetabolous). The aquatic larva or nymph hatches from an egg, and develops through eight to seventeen instars before leaving the water and emerging as the winged adult or imago.

Brachyopa flavescens, The Yellow Sapeater, is a fairly common species of syrphid fly. It has been observed in northeastern North America. Hoverflies get their names from the ability to remain nearly motionless while in flight. The adults are also known as flower flies for they are commonly found around and on flowers, from which they get both energy-giving nectar and protein-rich pollen. Larvae for this genus are of the rat-tailed type. B.flavescens larvae have not been described.
Blera nigra , the golden-haired wood fly, is a fairly common species of syrphid fly first officially described by Williston in 1887 Hoverflies get their name from the ability to remain nearly motionless while in flight The adults are also known as flower flies for they are commonly found around and on flowers from which they get both energy-giving nectar and protein rich pollen. The larvae are of the rat-tailed type feeding on exuding sap or in the rot holes of trees.

Blera robusta, the greenish wood fly, is a rare species of syrphid fly first officially described by Curran in 1922. Hoverflies get this name from the ability to remain nearly motionless while in flight. The adults are also known as flower flies, for they are commonly found around and on the flowers from which they get both energy-giving nectar and protein rich pollen. The larvae are of the rat-tailed type, feeding on exuding sap or in the rot holes of trees.

























{kind=link}