Related Research Articles

Meiosis is a special type of cell division of germ cells in sexually-reproducing organisms that produces the gametes, such as sperm or egg cells. It involves two rounds of division that ultimately result in four cells with only one copy of each chromosome (haploid). Additionally, prior to the division, genetic material from the paternal and maternal copies of each chromosome is crossed over, creating new combinations of code on each chromosome. Later on, during fertilisation, the haploid cells produced by meiosis from a male and female will fuse to create a cell with two copies of each chromosome again, the zygote.

Cell division is the process by which a parent cell divides, when a mother cell divides into two or more daughter cells. Cell division usually occurs as part of a larger cell cycle. In eukaryotes, there are two distinct types of cell division; a vegetative division, whereby each daughter cell is genetically identical to the parent cell (mitosis), and a reproductive cell division, whereby the number of chromosomes in the daughter cells is reduced by half to produce haploid gametes (meiosis). In cell biology, mitosis (/maɪˈtoʊsɪs/) is a part of the cell cycle, in which, replicated chromosomes are separated into two new nuclei. Cell division gives rise to genetically identical cells in which the total number of chromosomes is maintained. In general, mitosis is preceded by the S stage of interphase and is often followed by telophase and cytokinesis; which divides the cytoplasm, organelles, and cell membrane of one cell into two new cells containing roughly equal shares of these cellular components. The different stages of mitosis all together define the mitotic (M) phase of animal cell cycle—the division of the mother cell into two genetically identical daughter cells. Meiosis results in four haploid daughter cells by undergoing one round of DNA replication followed by two divisions. Homologous chromosomes are separated in the first division, and sister chromatids are separated in the second division. Both of these cell division cycles are used in the process of sexual reproduction at some point in their life cycle. Both are believed to be present in the last eukaryotic common ancestor.

Chromosomal crossover, or crossing over, is the exchange of genetic material during sexual reproduction between two homologous chromosomes' non-sister chromatids that results in recombinant chromosomes. It is one of the final phases of genetic recombination, which occurs in the pachytene stage of prophase I of meiosis during a process called synapsis. Synapsis begins before the synaptonemal complex develops and is not completed until near the end of prophase I. Crossover usually occurs when matching regions on matching chromosomes break and then reconnect to the other chromosome.
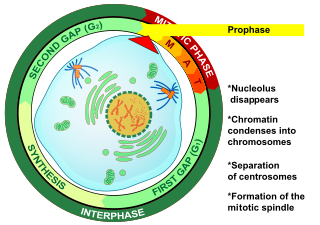
Prophase is the first stage of cell division in both mitosis and meiosis. Beginning after interphase, DNA has already been replicated when the cell enters prophase. The main occurrences in prophase are the condensation of the chromatin reticulum and the disappearance of the nucleolus.

Genetic recombination is the exchange of genetic material between different organisms which leads to production of offspring with combinations of traits that differ from those found in either parent. In eukaryotes, genetic recombination during meiosis can lead to a novel set of genetic information that can be further passed on from parents to offspring. Most recombination occurs naturally and can be classified into two types: (1) interchromosomal recombination, occurring through independent assortment of alleles whose loci are on different but homologous chromosomes ; & (2) intrachromosomal recombination, occurring through crossing over.

A unicellular organism, also known as a single-celled organism, is an organism that consists of a single cell, unlike a multicellular organism that consists of multiple cells. Organisms fall into two general categories: prokaryotic organisms and eukaryotic organisms. All prokaryotes are unicellular and are classified into bacteria and archaea. Many eukaryotes are multicellular, but some are unicellular such as protozoa, unicellular algae, and unicellular fungi. Unicellular organisms are thought to be the oldest form of life, with early protocells possibly emerging 3.8–4.0 billion years ago.

A couple of homologous chromosomes, or homologs, are a set of one maternal and one paternal chromosome that pair up with each other inside a cell during fertilization. Homologs have the same genes in the same loci where they provide points along each chromosome which enable a pair of chromosomes to align correctly with each other before separating during meiosis. This is the basis for Mendelian inheritance which characterizes inheritance patterns of genetic material from an organism to its offspring parent developmental cell at the given time and area.

In biology, a biological life cycle is a series of changes in form that an organism undergoes, returning to the starting state. "The concept is closely related to those of the life history, development and ontogeny, but differs from them in stressing renewal." Transitions of form may involve growth, asexual reproduction, or sexual reproduction.

Sexual reproduction is an adaptive feature which is common to almost all multicellular organisms and various unicellular organisms, with some organisms being incapable of asexual reproduction. Prior to the advent of sexual reproduction, the adaptation process whereby genes would change from one generation to the next happened very slowly and randomly. Sex evolved as an extremely efficient mechanism for producing variation, and this had the major advantage of enabling organisms to adapt to changing environments. Sex did, however, come with a cost. In reproducing asexually, no time nor energy needs to be expended in choosing a mate. And if the environment has not changed, then there may be little reason for variation, as the organism may already be well adapted. Sex also halves the amount of offspring a given population is able to produce. Sex, however, has evolved as the most prolific means of species branching into the tree of life. Diversification into the phylogenetic tree happens much more rapidly via sexual reproduction than it does by way of asexual reproduction.
RecQ helicase is a family of helicase enzymes initially found in Escherichia coli that has been shown to be important in genome maintenance. They function through catalyzing the reaction ATP + H2O → ADP + P and thus driving the unwinding of paired DNA and translocating in the 3' to 5' direction. These enzymes can also drive the reaction NTP + H2O → NDP + P to drive the unwinding of either DNA or RNA.

Homologous recombination is a type of genetic recombination in which genetic information is exchanged between two similar or identical molecules of double-stranded or single-stranded nucleic acids. It is widely used by cells to accurately repair harmful breaks that occur on both strands of DNA, known as double-strand breaks (DSB), in a process called homologous recombinational repair (HRR). Homologous recombination also produces new combinations of DNA sequences during meiosis, the process by which eukaryotes make gamete cells, like sperm and egg cells in animals. These new combinations of DNA represent genetic variation in offspring, which in turn enables populations to adapt during the course of evolution. Homologous recombination is also used in horizontal gene transfer to exchange genetic material between different strains and species of bacteria and viruses.

Synapsis is the pairing of two chromosomes that occurs during meiosis. It allows matching-up of homologous pairs prior to their segregation, and possible chromosomal crossover between them. Synapsis takes place during prophase I of meiosis. When homologous chromosomes synapse, their ends are first attached to the nuclear envelope. These end-membrane complexes then migrate, assisted by the extranuclear cytoskeleton, until matching ends have been paired. Then the intervening regions of the chromosome are brought together, and may be connected by a protein-RNA complex called the synaptonemal complex. During synapsis, autosomes are held together by the synaptonemal complex along their whole length, whereas for sex chromosomes, this only takes place at one end of each chromosome.
The yeast Saccharomyces cerevisiae is a simple single-celled eukaryote with both a diploid and haploid mode of existence. The mating of yeast only occurs between haploids, which can be either the a or α (alpha) mating type and thus display simple sexual differentiation. Mating type is determined by a single locus, MAT, which in turn governs the sexual behaviour of both haploid and diploid cells. Through a form of genetic recombination, haploid yeast can switch mating type as often as every cell cycle.

In microbiology, genetics, cell biology, and molecular biology, competence is the ability of a cell to alter its genetics by taking up extracellular ("naked") DNA from its environment in the process called transformation. Competence may be differentiated between natural competence, a genetically specified ability of bacteria which is thought to occur under natural conditions as well as in the laboratory, and induced or artificial competence, which arises when cells in laboratory cultures are treated to make them transiently permeable to DNA. Competence allows for rapid adaptation and DNA repair of the cell. This article primarily deals with natural competence in bacteria, although information about artificial competence is also provided.
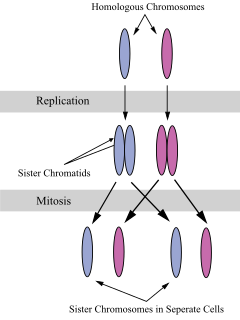
A sister chromatid refers to the identical copies (chromatids) formed by the DNA replication of a chromosome, with both copies joined together by a common centromere. In other words, a sister chromatid may also be said to be 'one-half' of the duplicated chromosome. A pair of sister chromatids is called a dyad. A full set of sister chromatids is created during the synthesis (S) phase of interphase, when all the chromosomes in a cell are replicated. The two sister chromatids are separated from each other into two different cells during mitosis or during the second division of meiosis.

Sister chromatid exchange (SCE) is the exchange of genetic material between two identical sister chromatids.
Recombinases are genetic recombination enzymes.
Microbial genetics is a subject area within microbiology and genetic engineering. Microbial genetics studies microorganisms for different purposes. The microorganisms that are observed are bacteria, and archaea. Some fungi and protozoa are also subjects used to study in this field. The studies of microorganisms involve studies of genotype and expression system. Genotypes are the inherited compositions of an organism. Genetic Engineering is a field of work and study within microbial genetics. The usage of recombinant DNA technology is a process of this work. The process involves creating recombinant DNA molecules through manipulating a DNA sequence. That DNA created is then in contact with a host organism. Cloning is also an example of genetic engineering.
Chromosome segregation is the process in eukaryotes by which two sister chromatids formed as a consequence of DNA replication, or paired homologous chromosomes, separate from each other and migrate to opposite poles of the nucleus. This segregation process occurs during both mitosis and meiosis. Chromosome segregation also occurs in prokaryotes. However, in contrast to eukaryotic chromosome segregation, replication and segregation are not temporally separated. Instead segregation occurs progressively following replication.

Sexual reproduction is a type of reproduction that involves a complex life cycle in which a gamete with a single set of chromosomes combines with another gamete to produce a zygote that develops into an organism composed of cells with two sets of chromosomes (diploid). This is typical in animals, though number of chromosome sets and how that number changes in sexual reproduction varies, especially among plants, fungi, and other eukaryotes.
References
- ↑ Retallack GJ, Krull ES, Thackeray GD, Parkinson D (2013-09-01). "Problematic urn-shaped fossils from a Paleoproterozoic (2.2 Ga) paleosol in South Africa". Precambrian Research. 235: 71–87. Bibcode:2013PreR..235...71R. doi:10.1016/j.precamres.2013.05.015.
- 1 2 3 4 5 Harris Bernstein, Carol Bernstein, Evolutionary Origin of Recombination during Meiosis, BioScience, Volume 60, Issue 7, July/August 2010, Pages 498–505, https://doi.org/10.1525/bio.2010.60.7.5
- 1 2 Bernstein H, Bernstein C (2017). "Sexual Communication in Archaea, the Precursor to Eukaryotic Meiosis". In Witzany G (ed.). Biocommunication of Archaea. Springer Nature. pp. 301–117. doi : 10.1007/978-3-319-65536-9_7. ISBN 978-3-319-65535-2.
- ↑ Wilkins AS, Holliday R (January 2009). "The evolution of meiosis from mitosis". Genetics. 181 (1): 3–12. doi:10.1534/genetics.108.099762. PMC 2621177 . PMID 19139151.
- ↑ Villeneuve AM, Hillers KJ (September 2001). "Whence meiosis?". Cell. 106 (6): 647–50. doi: 10.1016/s0092-8674(01)00500-1 . PMID 11572770. S2CID 9743579.
- ↑ Ramesh MA, Malik SB, Logsdon JM (January 2005). "A phylogenomic inventory of meiotic genes; evidence for sex in Giardia and an early eukaryotic origin of meiosis". Current Biology. 15 (2): 185–91. doi: 10.1016/j.cub.2005.01.003 . PMID 15668177. S2CID 17013247.
- ↑ Poxleitner MK, Carpenter ML, Mancuso JJ, Wang CJ, Dawson SC, Cande WZ (March 2008). "Evidence for karyogamy and exchange of genetic material in the binucleate intestinal parasite Giardia intestinalis". Science. 319 (5869): 1530–3. Bibcode:2008Sci...319.1530P. doi:10.1126/science.1153752. PMID 18339940. S2CID 206510785.
- ↑ Malik SB, Pightling AW, Stefaniak LM, Schurko AM, Logsdon JM (August 2007). Hahn MW (ed.). "An expanded inventory of conserved meiotic genes provides evidence for sex in Trichomonas vaginalis". PLOS ONE. 3 (8): e2879. Bibcode:2008PLoSO...3.2879M. doi: 10.1371/journal.pone.0002879 . PMC 2488364 . PMID 18663385.
- 1 2 3 Wilkins AS, Holliday R (January 2009). "The evolution of meiosis from mitosis". Genetics. 181 (1): 3–12. doi:10.1534/genetics.108.099762. PMC 2621177 . PMID 19139151.
- ↑ Herskowitz I (December 1988). "Life cycle of the budding yeast Saccharomyces cerevisiae". Microbiological Reviews. 52 (4): 536–53. doi:10.1128/MMBR.52.4.536-553.1988. PMC 373162 . PMID 3070323.
- ↑ Sager R, Granick S (July 1954). "Nutritional control of sexuality in Chlamydomonas reinhardi". The Journal of General Physiology. 37 (6): 729–42. doi:10.1085/jgp.37.6.729. PMC 2147466 . PMID 13174779.
- ↑ Bernstein C, Johns V (April 1989). "Sexual reproduction as a response to H2O2 damage in Schizosaccharomyces pombe". Journal of Bacteriology. 171 (4): 1893–7. doi:10.1128/jb.171.4.1893-1897.1989. PMC 209837 . PMID 2703462.
- ↑ Nedelcu AM, Michod RE (November 2003). "Sex as a response to oxidative stress: the effect of antioxidants on sexual induction in a facultatively sexual lineage". Proceedings: Biological Sciences. 270 Suppl 2 (Suppl 2): S136–9. doi:10.1098/rsbl.2003.0062. PMC 1809951 . PMID 14667362.
- ↑ Kirk DL, Kirk MM (January 1986). "Heat shock elicits production of sexual inducer in Volvox". Science. 231 (4733): 51–4. Bibcode:1986Sci...231...51K. doi:10.1126/science.3941891. PMID 3941891.
- ↑ Anagnostopoulos C, Spizizen J (May 1961). "Requirements for Transformation in Bacillus Subtilis". Journal of Bacteriology. 81 (5): 741–6. doi:10.1128/JB.81.5.741-746.1961. PMC 279084 . PMID 16561900.
- ↑ Goodgal SH, Herriott RM (July 1961). "Studies on transformations of Hemophilus influenzae. I. Competence". The Journal of General Physiology. 44 (6): 1201–27. doi:10.1085/jgp.44.6.1201. PMC 2195138 . PMID 13707010.
- ↑ Aspiras MB, Ellen RP, Cvitkovitch DG (September 2004). "ComX activity of Streptococcus mutans growing in biofilms". FEMS Microbiology Letters. 238 (1): 167–74. doi:10.1016/j.femsle.2004.07.032. PMID 15336418.
- ↑ Claverys JP, Prudhomme M, Martin B (2006). "Induction of competence regulons as a general response to stress in gram-positive bacteria". Annual Review of Microbiology. 60: 451–75. doi:10.1146/annurev.micro.60.080805.142139. PMID 16771651.
- ↑ Bernstein H, Byerly HC, Hopf FA, Michod RE (September 1985). "Genetic damage, mutation, and the evolution of sex". Science. 229 (4719): 1277–81. Bibcode:1985Sci...229.1277B. doi:10.1126/science.3898363. PMID 3898363.
- ↑ Michod RE, Bernstein H, Nedelcu AM (May 2008). "Adaptive value of sex in microbial pathogens". Infection, Genetics and Evolution. 8 (3): 267–85. doi:10.1016/j.meegid.2008.01.002. PMID 18295550. http://www.hummingbirds.arizona.edu/Faculty/Michod/Downloads/IGE%20review%20sex.pdf
- ↑ Vilenchik MM, Knudson AG (October 2003). "Endogenous DNA double-strand breaks: production, fidelity of repair, and induction of cancer". Proceedings of the National Academy of Sciences of the United States of America. 100 (22): 12871–6. Bibcode:2003PNAS..10012871V. doi: 10.1073/pnas.2135498100 . PMC 240711 . PMID 14566050.
- ↑ Lenormand T, Engelstädter J, Johnston SE, Wijnker E, Haag CR (October 2016). "Evolutionary mysteries in meiosis". Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 371 (1706): 050831. bioRxiv 10.1101/050831 . doi: 10.1098/rstb.2016.0001 . PMC 5031626 . PMID 27619705.
- ↑ Marcon E, Moens PB (August 2005). "The evolution of meiosis: recruitment and modification of somatic DNA-repair proteins". BioEssays. 27 (8): 795–808. doi:10.1002/bies.20264. PMID 16015600. S2CID 27658497.
- 1 2 Hörandl E, Hadacek F (December 2013). "The oxidative damage initiation hypothesis for meiosis". Plant Reproduction. 26 (4): 351–67. doi:10.1007/s00497-013-0234-7. PMC 3825497 . PMID 23995700.
- ↑ Bernstein H, Bernstein C, Michod RE (2012). "DNA repair as the primary adaptive function of sex in bacteria and eukaryotes". International Journal of Medical and Biological Frontiers. 18. ISSN 1081-3829.
- 1 2 3 Mira A (September 1998). "Why is meiosis arrested?". Journal of Theoretical Biology. 194 (2): 275–87. doi:10.1006/jtbi.1998.0761. PMID 9778439.
- 1 2 3 Stringer JM, Winship A, Zerafa N, Wakefield M, Hutt K (May 2020). "Oocytes can efficiently repair DNA double-strand breaks to restore genetic integrity and protect offspring health". Proceedings of the National Academy of Sciences of the United States of America. 117 (21): 11513–11522. doi:10.1073/pnas.2001124117. PMC 7260990 . PMID 32381741.
- ↑ Otto SP, Gerstein AC (August 2006). "Why have sex? The population genetics of sex and recombination". Biochemical Society Transactions. 34 (Pt 4): 519–22. doi:10.1042/BST0340519. PMID 16856849.
