
Merostomata is a class of chelicerate arthropods that contains the extinct Eurypterida and the extant Xiphosura. The term was originally used by James Dwight Dana to refer to Xiphosura only, but was emended by Henry Woodward to cover both groups.

The subphylum Chelicerata constitutes one of the major subdivisions of the phylum Arthropoda. Chelicerates include the sea spiders, horseshoe crabs, and arachnids, as well as a number of extinct lineages, such as the eurypterids and chasmataspidids.

Arachnida is a class of joint-legged arthropods, in the subphylum Chelicerata. Arachnida includes, among others, spiders, scorpions, ticks, mites, pseudoscorpions, harvestmen, camel spiders, whip spiders and vinegaroons.

Ricinulei is a small order of arachnids. Like most arachnids, they are predatory, eating small arthropods. They occur today in west-central Africa (Ricinoides) and the Americas as far north as Texas. As of 2021, 91 extant species of ricinuleids have been described worldwide, all in the single family Ricinoididae. In older works they are sometimes referred to as Podogona. Due to their obscurity they do not have a proper common name, though in academic literature they are occasionally referred to as hooded tickspiders.

The Mesothelae are a suborder of spiders that includes a single extant family, Liphistiidae, and a number of extinct families. This suborder is thought to form the sister group to all other living spiders, and to retain ancestral characters, such as a segmented abdomen with spinnerets in the middle and two pairs of book lungs. Members of Liphistiidae are medium to large spiders with eight eyes grouped on a tubercle. They are found only in China, Japan, and southeast Asia. The oldest known Mesothelae spiders are known from the Carboniferous, over 300 million years ago.

Xiphosura is an order of arthropods related to arachnids. They are more commonly known as horseshoe crabs. They first appeared in the Hirnantian. Currently, there are only four living species. Xiphosura contains one suborder, Xiphosurida, and several stem-genera.

Plesiosiro is an extinct arachnid genus known exclusively from nine specimens from the Upper Carboniferous of Coseley, Staffordshire, United Kingdom. The genus is monotypic, represented only by the species Plesiosiro madeleyi described by Reginald Innes Pocock in his important 1911 monograph on British Carboniferous arachnids. It is the only known member of the order Haptopoda.
The order Trigonotarbida is a group of extinct arachnids whose fossil record extends from the late Silurian to the early Permian. These animals are known from several localities in Europe and North America, as well as a single record from Argentina. Trigonotarbids can be envisaged as spider-like arachnids, but without silk-producing spinnerets. They ranged in size from a few millimetres to a few centimetres in body length and had segmented abdomens (opisthosoma), with the dorsal exoskeleton (tergites) across the backs of the animals' abdomens, which were characteristically divided into three or five separate plates. Probably living as predators on other arthropods, some later trigonotarbid species were quite heavily armoured and protected themselves with spines and tubercles. About seventy species are currently known, with most fossils originating from the Carboniferous coal measures.

Tetrapulmonata is a non-ranked supra-ordinal clade of arachnids. It is composed of the extant orders Uropygi, Schizomida, Amblypygi and Araneae (spiders). It is the only supra-ordinal group of arachnids that is strongly supported in molecular phylogenetic studies. Two extinct orders are also placed in this clade, Haptopoda and Uraraneida. In 2016, a newly described fossil arachnid, Idmonarachne, was also included in the Tetrapulmonata; as of March 2016 it has not been assigned to an order.
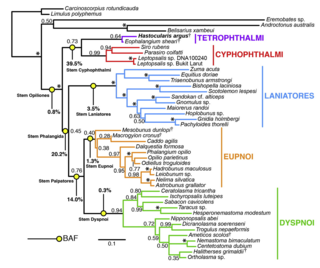
Harvestmen (Opiliones) are an order of arachnids often confused with spiders, though the two orders are not closely related. Research on harvestman phylogeny is in a state of flux. While some families are clearly monophyletic, that is share a common ancestor, others are not, and the relationships between families are often not well understood.
Attercopus is an extinct genus of arachnids, containing one species Attercopus fimbriunguis, known from flattened cuticle fossils from the Panther Mountain Formation in Upstate New York. It is placed in the extinct order Uraraneida, spider-like animals able to produce silk, but which lacked true spinnerets and retained a segmented abdomen bearing a flagellum-like tail resembling that of a whip scorpion. They are thought to be close to the origins of spiders.

Eophrynus prestvicii is an extinct species of arachnid belonging to the order Trigonotarbida.

Eophrynidae is a family of the extinct arachnid order Trigonotarbida. Eophrynids lived during the Carboniferous period in what is now modern Europe and North America. The family is probably found within the "eophrynid assemblage" clade:.

Spiders are air-breathing arthropods that have eight limbs, chelicerae with fangs generally able to inject venom, and spinnerets that extrude silk. They are the largest order of arachnids and rank seventh in total species diversity among all orders of organisms. Spiders are found worldwide on every continent except for Antarctica, and have become established in nearly every land habitat. As of November 2023, 51,673 spider species in 136 families have been recorded by taxonomists. However, there has been debate among scientists about how families should be classified, with over 20 different classifications proposed since 1900.

Palaeocharinus is a genus of extinct trigonotarbid arachnids known from the Devonian of western Europe. The genus was first found and described in the Rhynie chert in the 1920s by Arthur Stanley Hirst and S. Maulik. The family to which the genus belongs may be paraphyletic.
Rosamygale is a genus of extinct Triassic spiders, with a single described species, Rosamygale grauvogeli. It is the oldest known member of the Mygalomorphae, one of the three main divisions of spiders, which includes well known forms such as tarantulas and Australian funnel-web spiders. It was described by Selden and Gall in 1992, from specimens found in the Middle Triassic aged Gres a Meules and Grès à Voltzia geological formations in France. It is also considered to be the oldest known member of the Avicularioidea, one of the two main divisions of Mygalomorphae.
Uraraneida is an extinct order of Paleozoic arachnids related to modern spiders. Two genera of fossils have been definitively placed in this order: Attercopus from the Devonian of United States and Permarachne from the Permian of Russia. Like spiders, they are known to have produced silk, but lack the characteristic spinnerets of modern spiders, and retain elongate telsons.

Idmonarachne is an extinct genus of arachnids, containing one species, Idmonarachne brasieri. It is related to uraraneids and spiders.
This list of fossil arthropods described in 2011 is a list of new taxa of trilobites, fossil insects, crustaceans, arachnids and other fossil arthropods of every kind that have been described during the year 2011. The list only includes taxa at the level of genus or species.

Chimerarachne is a genus of extinct arachnids, containing five species. Fossils of Chimerarachne were discovered in Burmese amber from Myanmar which dates to the mid-Cretaceous, about 100 million years ago. It is thought to be closely related to spiders, but outside any living spider clade. The earliest spider fossils are from the Carboniferous, requiring at least a 170 myr ghost lineage with no fossil record. The size of the animal is quite small, being only 2.5 millimetres (0.098 in) in body length, with the tail being about 3 millimetres (0.12 in) in length. These fossils resemble spiders in having two of their key defining features: spinnerets for spinning silk, and a modified male organ on the pedipalp for transferring sperm. At the same time they retain a whip-like tail, rather like that of a whip scorpion and uraraneids. Chimerarachne is not ancestral to spiders, being much younger than the oldest spiders which are known from the Carboniferous, but it appears to be a late survivor of an extinct group which was probably very close to the origins of spiders. It suggests that there used to be spider-like animals with tails which lived alongside true spiders for at least 200 million years.


