
Evolution is the change in the heritable characteristics of biological populations over successive generations. It occurs when evolutionary processes such as natural selection and genetic drift act on genetic variation, resulting in certain characteristics becoming more or less common within a population over successive generations. The process of evolution has given rise to biodiversity at every level of biological organisation.
Evolutionary psychology is a theoretical approach in psychology that examines cognition and behavior from a modern evolutionary perspective. It seeks to identify human psychological adaptations with regards to the ancestral problems they evolved to solve. In this framework, psychological traits and mechanisms are either functional products of natural and sexual selection or non-adaptive by-products of other adaptive traits.
Microevolution is the change in allele frequencies that occurs over time within a population. This change is due to four different processes: mutation, selection, gene flow and genetic drift. This change happens over a relatively short amount of time compared to the changes termed macroevolution.

Natural selection is the differential survival and reproduction of individuals due to differences in phenotype. It is a key mechanism of evolution, the change in the heritable traits characteristic of a population over generations. Charles Darwin popularised the term "natural selection", contrasting it with artificial selection, which is intentional, whereas natural selection is not.

Sexual selection is a mode of natural selection in which members of one biological sex choose mates of the other sex to mate with, and compete with members of the same sex for access to members of the opposite sex. These two forms of selection mean that some individuals have greater reproductive success than others within a population, for example because they are more attractive or prefer more attractive partners to produce offspring. Successful males benefit from frequent mating and monopolizing access to one or more fertile females. Females can maximise the return on the energy they invest in reproduction by selecting and mating with the best males.

Convergent evolution is the independent evolution of similar features in species of different periods or epochs in time. Convergent evolution creates analogous structures that have similar form or function but were not present in the last common ancestor of those groups. The cladistic term for the same phenomenon is homoplasy. The recurrent evolution of flight is a classic example, as flying insects, birds, pterosaurs, and bats have independently evolved the useful capacity of flight. Functionally similar features that have arisen through convergent evolution are analogous, whereas homologous structures or traits have a common origin but can have dissimilar functions. Bird, bat, and pterosaur wings are analogous structures, but their forelimbs are homologous, sharing an ancestral state despite serving different functions.

Vestigiality is the retention, during the process of evolution, of genetically determined structures or attributes that have lost some or all of the ancestral function in a given species. Assessment of the vestigiality must generally rely on comparison with homologous features in related species. The emergence of vestigiality occurs by normal evolutionary processes, typically by loss of function of a feature that is no longer subject to positive selection pressures when it loses its value in a changing environment. The feature may be selected against more urgently when its function becomes definitively harmful, but if the lack of the feature provides no advantage, and its presence provides no disadvantage, the feature may not be phased out by natural selection and persist across species.

This is a list of topics in evolutionary biology.
Experimental evolution is the use of laboratory experiments or controlled field manipulations to explore evolutionary dynamics. Evolution may be observed in the laboratory as individuals/populations adapt to new environmental conditions by natural selection.
Evolvability is defined as the capacity of a system for adaptive evolution. Evolvability is the ability of a population of organisms to not merely generate genetic diversity, but to generate adaptive genetic diversity, and thereby evolve through natural selection.
In biology, adaptation has three related meanings. Firstly, it is the dynamic evolutionary process of natural selection that fits organisms to their environment, enhancing their evolutionary fitness. Secondly, it is a state reached by the population during that process. Thirdly, it is a phenotypic trait or adaptive trait, with a functional role in each individual organism, that is maintained and has evolved through natural selection.

A unit of selection is a biological entity within the hierarchy of biological organization that is subject to natural selection. There is debate among evolutionary biologists about the extent to which evolution has been shaped by selective pressures acting at these different levels.
Adaptationism is a scientific perspective on evolution that focuses on accounting for the products of evolution as collections of adaptive traits, each a product of natural selection with some adaptive rationale or raison d'etre. A formal alternative would be to look at the products of evolution as the result of neutral evolution, in terms of structural constraints, or in terms of a mixture of factors including natural selection.

Divergent evolution or divergent selection is the accumulation of differences between closely related populations within a species, sometimes leading to speciation. Divergent evolution is typically exhibited when two populations become separated by a geographic barrier and experience different selective pressures that cause adaptations. After many generations and continual evolution, the populations become less able to interbreed with one another. The American naturalist J. T. Gulick (1832–1923) was the first to use the term "divergent evolution", with its use becoming widespread in modern evolutionary literature. Examples of divergence in nature are the adaptive radiation of the finches of the Galápagos, changes in mobbing behavior of the kittiwake, and the evolution of the modern-day dog from the wolf.

Elisabeth S. Vrba is a paleontologist at Yale University who developed the turnover-pulse hypothesis.
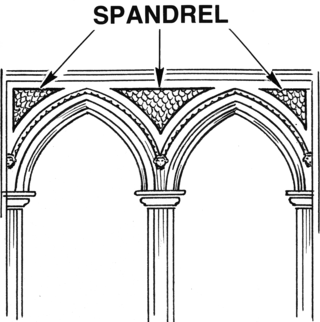
In evolutionary biology, a spandrel is a phenotypic trait that is a byproduct of the evolution of some other characteristic, rather than a direct product of adaptive selection. Stephen Jay Gould and Richard Lewontin brought the term into biology in their 1979 paper "The Spandrels of San Marco and the Panglossian Paradigm: A Critique of the Adaptationist Programme". Adaptationism is a point of view that sees most organismal traits as adaptive products of natural selection. Gould and Lewontin sought to temper what they saw as adaptationist bias by promoting a more structuralist view of evolution.
Evolutionary psychology seeks to identify and understand human psychological traits that have evolved in much the same way as biological traits, through adaptation to environmental cues. Furthermore, it tends toward viewing the vast majority of psychological traits, certainly the most important ones, as the result of past adaptions, which has generated significant controversy and criticism from competing fields. These criticisms include disputes about the testability of evolutionary hypotheses, cognitive assumptions such as massive modularity, vagueness stemming from assumptions about the environment that leads to evolutionary adaptation, the importance of non-genetic and non-adaptive explanations, as well as political and ethical issues in the field itself.

Evolutionary thought, the recognition that species change over time and the perceived understanding of how such processes work, has roots in antiquity—in the ideas of the ancient Greeks, Romans, Chinese, Church Fathers as well as in medieval Islamic science. With the beginnings of modern biological taxonomy in the late 17th century, two opposed ideas influenced Western biological thinking: essentialism, the belief that every species has essential characteristics that are unalterable, a concept which had developed from medieval Aristotelian metaphysics, and that fit well with natural theology; and the development of the new anti-Aristotelian approach to modern science: as the Enlightenment progressed, evolutionary cosmology and the mechanical philosophy spread from the physical sciences to natural history. Naturalists began to focus on the variability of species; the emergence of palaeontology with the concept of extinction further undermined static views of nature. In the early 19th century prior to Darwinism, Jean-Baptiste Lamarck (1744–1829) proposed his theory of the transmutation of species, the first fully formed theory of evolution.
Evolutionary psychology of language is the study of the evolutionary history of language as a psychological faculty within the discipline of evolutionary psychology. It makes the assumption that language is the result of a Darwinian adaptation.

Teleology in biology is the use of the language of goal-directedness in accounts of evolutionary adaptation, which some biologists and philosophers of science find problematic. The term teleonomy has also been proposed. Before Darwin, organisms were seen as existing because God had designed and created them; their features such as eyes were taken by natural theology to have been made to enable them to carry out their functions, such as seeing. Evolutionary biologists often use similar teleological formulations that invoke purpose, but these imply natural selection rather than actual goals, whether conscious or not. Some biologists and religious thinkers held that evolution itself was somehow goal-directed (orthogenesis), and in vitalist versions, driven by a purposeful life force. With evolution working by natural selection acting on inherited variation, the use of teleology in biology has attracted criticism, and attempts have been made to teach students to avoid teleological language.

