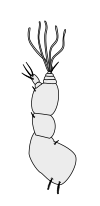
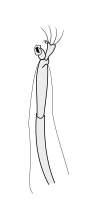
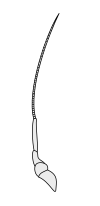
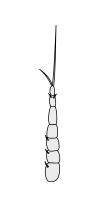
Halteres are a pair of small club-shaped organs on the body of two orders of flying insects that provide information about body rotations during flight. Insects of the large order Diptera (flies) have halteres which evolved from a pair of ancestral hindwings, while males of the much smaller order Strepsiptera (stylops) have halteres which evolved from a pair of ancestral forewings.

Dendrobranchiata is a suborder of decapods, commonly known as prawns. There are 540 extant species in seven families, and a fossil record extending back to the Devonian. They differ from related animals, such as Caridea and Stenopodidea, by the branching form of the gills and by the fact that they do not brood their eggs, but release them directly into the water. They may reach a length of over 330 millimetres (13 in) and a mass of 450 grams (1.0 lb), and are widely fished and farmed for human consumption.

Malacostraca is the second largest of the six classes of pancrustaceans just behind hexapods, containing about 40,000 living species, divided among 16 orders. Its members, the malacostracans, display a great diversity of body forms and include crabs, lobsters, crayfish, shrimp, krill, prawns, woodlice, amphipods, mantis shrimp, tongue-eating lice and many other less familiar animals. They are abundant in all marine environments and have colonised freshwater and terrestrial habitats. They are segmented animals, united by a common body plan comprising 20 body segments, and divided into a head, thorax, and abdomen.

An appendage is an external body part, or natural prolongation, that protrudes from an organism's or microorganism's body.

Johnston's organ is a collection of sensory cells found in the pedicel of the antennae in the class Insecta. Johnston's organ detects motion in the flagellum. It consists of scolopidia arrayed in a bowl shape, each of which contains a mechanosensory chordotonal neuron. The number of scolopidia varies between species. In homopterans, the Johnston's organs contain 25 - 79 scolopidia. The presence of Johnston's organ is a defining characteristic which separates the class Insecta from the other hexapods belonging to the group Entognatha. Johnston's organ was named after the physician Christopher Johnston (1822-1891) father of the physician and Assyriologist Christopher Johnston.

Canadaspis is an extinct genus of bivalved Cambrian arthropod, known from North America and China. They are thought to have been benthic feeders that moved mainly by walking and possibly used its biramous appendages to stir mud in search of food. They have been placed within the Hymenocarina, which includes other bivalved Cambrian arthropods.
The arthropod leg is a form of jointed appendage of arthropods, usually used for walking. Many of the terms used for arthropod leg segments are of Latin origin, and may be confused with terms for bones: coxa, trochanter, femur, tibia, tarsus, ischium, metatarsus, carpus, dactylus, patella.

This glossary of entomology describes terms used in the formal study of insect species by entomologists.

The Trichogrammatidae are a family of small endoparasitoid wasps in the superfamily Chalcidoidea that include some of the smallest of all insects, with most species having adults less than 1 mm in length, with species of Megaphragma having an adult body length less than 300 μm. Over 840 species are placed in about 80 genera; their distribution is worldwide.
Chordotonal organs are stretch receptor organs found only in insects and crustaceans. They are located at most joints and are made up of clusters of scolopidia that either directly or indirectly connect two joints and sense their movements relative to one another. They can have both extero- and proprioceptive functions, for example sensing auditory stimuli or leg movement. The word was coined by Vitus Graber in 1882, though he interpreted them as being stretched between two points like a string, sensing vibrations through resonance.

The (pan)arthropod head problem is a long-standing zoological dispute concerning the segmental composition of the heads of the various arthropod groups, and how they are evolutionarily related to each other. While the dispute has historically centered on the exact make-up of the insect head, it has been widened to include other living arthropods, such as chelicerates, myriapods, and crustaceans, as well as fossil forms, such as the many arthropods known from exceptionally preserved Cambrian faunas. While the topic has classically been based on insect embryology, in recent years a great deal of developmental molecular data has become available. Dozens of more or less distinct solutions to the problem, dating back to at least 1897, have been published, including several in the 2000s.

The subphylum Hexapoda or hexapods comprises the largest clade of arthropods and includes most of the extant arthropod species. It includes the crown group class Insecta, as well as the much smaller class Entognatha, which includes three orders of wingless arthropods that were once considered insects: Collembola (springtails), Protura (coneheads) and Diplura. The insects and springtails are very abundant and are some of the most important pollinators, basal consumers, scavengers/detritivores and micropredators in terrestrial environments.

Zigrasimecia is an extinct genus of ants which existed in the Cretaceous period approximately 98 million years ago. The first specimens were collected from Burmese amber in Kachin State, 100 kilometres (62 mi) west of Myitkyina town in Myanmar. In 2013, palaeoentomologists Phillip Barden and David Grimaldi published a paper describing and naming Zigrasimecia tonsora. They described a dealate female with unusual features, notably the highly specialized mandibles. Other features include large ocelli, short scapes, 12 antennomeres, small eyes, and a clypeal margin that has a row of peg-like denticles. The genus Zigrasimecia was originally incertae sedis within Formicidae until a second species, Zigrasimecia ferox, was described in 2014, leading to its placement in the subfamily Sphecomyrminae. Later, it was considered to belong to the distinct subfamily Zigrasimeciinae.

Phoracantha semipunctata, the Australian Eucalyptus longhorn, is a species of beetle in the family Cerambycidae. Native to Australia, it has now spread to many parts of the world, including practically all countries where tree species of Eucalyptus have been introduced. It has been classified as an invasive pest species of Eucalyptus outside Australia.

Insect olfaction refers to the function of chemical receptors that enable insects to detect and identify volatile compounds for foraging, predator avoidance, finding mating partners and locating oviposition habitats. Thus, it is the most important sensation for insects. Most important insect behaviors must be timed perfectly which is dependent on what they smell and when they smell it. For example, olfaction is essential for locating host plants and hunting prey in many species of insects, such as the moth Deilephila elpenor and the wasp Polybia sericea, respectively.
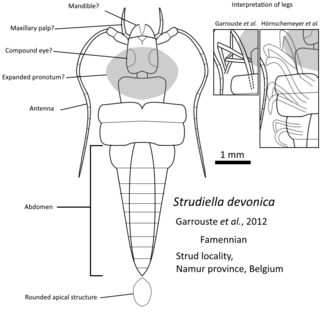
Strudiella devonica is a species of extinct arthropod from the Devonian. It was recovered in the Strud environment from the Bois des Mouches Formation, Upper Famennian. It was originally described as the first complete Late Devonian terrestrial insect, but due to its poor state of preservation, its affinity is discussed.

'Blera confusa , the confusing wood fly, is a common species of syrphid fly first officially described by Johnson, 1913 Hoverflies get their names from the ability to remain nearly motionless while in flight. The adults are also known as flower flies for they are commonly found around and on flowers, from which they get both energy-giving nectar and protein-rich pollen. The larvae are of the rat-tailed type feeding on exuding sap or in the rot holes of trees.
Polytus mellerborgi, commonly known as small banana weevil, is a species of weevil widely distributed in southeastern Polynesia through Melanesia, Micronesia, Indo-Malaya, south China, Burma, India, Sri Lanka, other islands of the Indian Ocean to Madagascar and as far as Mexico and Central America.

Bundenbachiellus is an extinct genus of arthropod described from the Lower Devonian Hunsrück Slate of Germany. This genus is known from only one species, B. giganteus. Alongside its possible relative Enalikter from Silurian, it is possible that genus is late-living example of Megacheira, "great-appendage arthropod".
Martinssonia is an extinct genus of Cambrian arthropod from the Orsten Lagerstätte.