
Biostratigraphy is the branch of stratigraphy which focuses on correlating and assigning relative ages of rock strata by using the fossil assemblages contained within them. The primary objective of biostratigraphy is correlation, demonstrating that a particular horizon in one geological section represents the same period of time as another horizon at a different section. Fossils within these strata are useful because sediments of the same age can look completely different, due to local variations in the sedimentary environment. For example, one section might have been made up of clays and marls, while another has more chalky limestones. However, if the fossil species recorded are similar, the two sediments are likely to have been laid down around the same time. Ideally these fossils are used to help identify biozones, as they make up the basic biostratigraphy units, and define geological time periods based upon the fossil species found within each section.

In the geologic timescale, the Anisian is the lower stage or earliest age of the Middle Triassic series or epoch and lasted from 247.2 million years ago until 242 million years ago. The Anisian Age succeeds the Olenekian Age and precedes the Ladinian Age.
The Messinian is in the geologic timescale the last age or uppermost stage of the Miocene. It spans the time between 7.246 ± 0.005 Ma and 5.333 ± 0.005 Ma. It follows the Tortonian and is followed by the Zanclean, the first age of the Pliocene.

The Carnian is the lowermost stage of the Upper Triassic Series. It lasted from 237 to 227 million years ago (Ma). The Carnian is preceded by the Ladinian and is followed by the Norian. Its boundaries are not characterized by major extinctions or biotic turnovers, but a climatic event occurred during the Carnian and seems to be associated with important extinctions or biotic radiations. Another extinction occurred at the Carnian-Norian boundary, ending the Carnian age.

The Ladinian is a stage and age in the Middle Triassic series or epoch. It spans the time between 242 Ma and ~237 Ma. The Ladinian was preceded by the Anisian and succeeded by the Carnian.
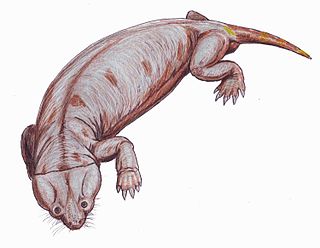
The Cistecephalus Assemblage Zone is a tetrapod assemblage zone or biozone found in the Adelaide Subgroup of the Beaufort Group, a majorly fossiliferous and geologically important geological group of the Karoo Supergroup in South Africa. This biozone has outcrops located in the Teekloof Formation north-west of Beaufort West in the Western Cape, in the upper Middleton and lower Balfour Formations respectively from Colesberg of the Northern Cape to east of Graaff-Reinet in the Eastern Cape. The Cistecephalus Assemblage Zone is one of eight biozones found in the Beaufort Group, and is considered to be Late Permian in age.
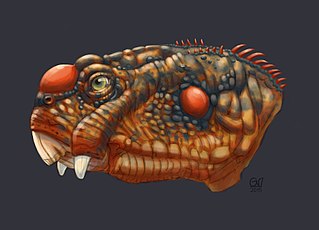
The Daptocephalus Assemblage Zone is a tetrapod assemblage zone or biozone found in the Adelaide Subgroup of the Beaufort Group, a majorly fossiliferous and geologically important Group of the Karoo Supergroup in South Africa. This biozone has outcrops located in the upper Teekloof Formation west of 24°E, the majority of the Balfour Formation east of 24°E, and the Normandien Formation in the north. It has numerous localities which are spread out from Colesberg in the Northern Cape, Graaff-Reniet to Mthatha in the Eastern Cape, and from Bloemfontein to Harrismith in the Free State. The Daptocephalus Assemblage Zone is one of eight biozones found in the Beaufort Group and is considered Late Permian (Lopingian) in age. Its contact with the overlying Lystrosaurus Assemblage Zone marks the Permian-Triassic boundary.

The Cynognathus Assemblage Zone is a tetrapod biozone utilized in the Karoo Basin of South Africa. It is equivalent to the Burgersdorp Formation, the youngest lithostratigraphic formation in the Beaufort Group, which is part of the fossiliferous and geologically important Karoo Supergroup. The Cynognathus Assemblage Zone is the youngest of the eight biozones found in the Beaufort Group, and is considered to be late Early Triassic (Olenekian) to early Middle Triassic (Anisian) in age. The name of the biozone refers to Cynognathus crateronotus, a large and carnivorous cynodont therapsid which occurs throughout the entire biozone.

The Pristerognathus Assemblage Zone is a tetrapod assemblage zone or biozone which correlates to the upper Abrahamskraal Formation and lowermost Teekloof Formation, Adelaide Subgroup of the Beaufort Group, a fossiliferous and geologically important geological Group of the Karoo Supergroup in South Africa. The thickest outcrops, reaching not more than 300 metres (980 ft), occur just east of Sutherland through to Beaufort West in the south and Victoria West in the north. Exposures are also found west of Colesberg and south of Graaff-Reinet. The Pristerognathus Assemblage Zone is the third biozone of the Beaufort Group.

The Dockum is a Late Triassic geologic group found primarily on the Llano Estacado of western Texas and eastern New Mexico with minor exposures in southwestern Kansas, eastern Colorado, and Oklahoma panhandle. The Dockum reaches a maximum thickness of slightly over 650 m but is usually much thinner. The Dockum rests on an unconformity over the Anisian aged Anton Chico Formation.
The Blancan North American Stage on the geologic timescale is the North American faunal stage according to the North American Land Mammal Ages chronology (NALMA), typically set from 4,750,000 to 1,806,000 years BP, a period of 2.944 million years. It is usually considered to start in the early-mid Pliocene Epoch and end by the early Pleistocene. The Blancan is preceded by the Hemphillian and followed by the Irvingtonian NALMA stages.
Redondasaurus is an extinct genus or subgenus of phytosaur from the Late Triassic of the southwestern United States. It was named by Hunt & Lucas in 1993, and contains two species, R. gregorii and R. bermani. It is the youngest and most evolutionarily-advanced of the phytosaurs.
The North American land mammal ages (NALMA) establishes a geologic timescale for North American fauna beginning during the Late Cretaceous and continuing through to the present. These periods are referred to as ages or intervals and were established using geographic place names where fossil materials were obtained.
The Irvingtonian North American Land Mammal Age on the geologic timescale is the North American faunal stage according to the North American Land Mammal Ages chronology (NALMA), spanning from 1.8 million – 250,000 years BP. Named after an assemblage of fossils from the Irvington District of Fremont, California, the Irvingtonian is usually considered to overlap the Lower Pleistocene and Middle Pleistocene epochs. The Irvingtonian is preceded by the Blancan and followed by the Rancholabrean NALMA stages.
The European Land Mammal Mega Zones are zones in rock layers that have a specific assemblage of fossils (biozones) based on occurrences of fossil assemblages of European land mammals. These biozones cover most of the Cenozoic, with particular focus having been paid to the Neogene and Paleogene systems, the Quaternary has several competing systems. In cases when fossils of mammals are abundant, stratigraphers and paleontologists can use these biozones as a more practical regional alternative to the stages of the official ICS geologic timescale. European Land Mammal Mega Zones are often also confusingly referred to as ages, stages, or intervals.
The Williams Fork Formation is a Campanian to Maastrichtian (Edmontonian) geologic formation of the Mesaverde Group in Colorado. Dinosaur remains are among the fossils that have been recovered from the formation, most notably Pentaceratops sternbergii,. Other fossils found in the formation are the ammonite Lewyites, tyrannosaurids, dromaeosaurids, troodontids, nodosaurids, ankylosaurids, hadrosaurids, hybodonts, neosuchian crocodylomorphs, and the mammals Glasbius and Meniscoessus collomensis.
The Norian is a division of the Triassic Period. It has the rank of an age (geochronology) or stage (chronostratigraphy). It lasted from ~227 to 208.5 million years ago. It was preceded by the Carnian and succeeded by the Rhaetian.

The Manda Formation is a Middle Triassic (Anisian?) or possibly Late Triassic (Carnian?) geologic formation in Tanzania. It preserves fossils of many terrestrial vertebrates from the Triassic, including some of the earliest dinosauromorph archosaurs. The formation is often considered to be Anisian in age according to general tetrapod biochronology hypotheses and correlations to the Cynognathus Assemblage Zone of South Africa. However, some recent studies cast doubt to this age, suggesting that parts deposits may actually be younger (Carnian) in age.

The Bone Valley Formation is a geologic formation in Florida. It is sometimes classified as the upper member of the Peace River Formation of the Hawthorn Group. It contains economically important phosphorite deposits that are mined in west-central Florida, as well as rich assemblages of vertebrate fossils.
Land vertebrate faunachrons (LVFs) are biochronological units used to correlate and date terrestrial sediments and fossils based on their tetrapod faunas. First formulated on a global scale by Spencer G. Lucas in 1998, LVFs are primarily used within the Triassic Period, though Lucas later designated LVFs for other periods as well. Eight worldwide LVFs are defined for the Triassic. The first two earliest Triassic LVFs, the Lootsbergian and Nonesian, are based on South African synapsids and faunal assemblage zones estimated to correspond to the Early Triassic. These are followed by the Perovkan and Berdyankian, based on temnospondyl amphibians and Russian assemblages estimated to be from the Middle Triassic. The youngest four Triassic LVFs, the Otischalkian, Adamanian, Revueltian, and Apachean, are based on aetosaur and phytosaur reptiles common in the Late Triassic of the southwestern United States.

