Related Research Articles

An endosymbiont or endobiont is any organism that lives within the body or cells of another organism most often, though not always, in a mutualistic relationship. This phenomenon is known as endosymbiosis. Examples are nitrogen-fixing bacteria, which live in the root nodules of legumes, single-cell algae inside reef-building corals and bacterial endosymbionts that provide essential nutrients to insects.

A mycorrhiza is a symbiotic association between a fungus and a plant. The term mycorrhiza refers to the role of the fungus in the plant's rhizosphere, its root system. Mycorrhizae play important roles in plant nutrition, soil biology, and soil chemistry.

An arbuscular mycorrhiza (AM) is a type of mycorrhiza in which the symbiont fungus penetrates the cortical cells of the roots of a vascular plant forming arbuscules. Arbuscular mycorrhiza is a type of endomycorrhiza along with ericoid mycorrhiza and orchid mycorrhiza .They are characterized by the formation of unique tree-like structures, the arbuscules. In addition, globular storage structures called vesicles are often encountered.

Root hair, or absorbent hairs, are outgrowths of epidermal cells, specialized cells at the tip of a plant root. They are lateral extensions of a single cell and are only rarely branched. They are found in the region of maturation, of the root. Root hair cells improve plant water absorption by increasing root surface area to volume ratio which allows the root hair cell to take in more water. The large vacuole inside root hair cells makes this intake much more efficient. Root hairs are also important for nutrient uptake as they are main interface between plants and mycorrhizal fungi.
Glomeromycota are one of eight currently recognized divisions within the kingdom Fungi, with approximately 230 described species. Members of the Glomeromycota form arbuscular mycorrhizas (AMs) with the thalli of bryophytes and the roots of vascular land plants. Not all species have been shown to form AMs, and one, Geosiphon pyriformis, is known not to do so. Instead, it forms an endocytobiotic association with Nostoc cyanobacteria. The majority of evidence shows that the Glomeromycota are dependent on land plants for carbon and energy, but there is recent circumstantial evidence that some species may be able to lead an independent existence. The arbuscular mycorrhizal species are terrestrial and widely distributed in soils worldwide where they form symbioses with the roots of the majority of plant species (>80%). They can also be found in wetlands, including salt-marshes, and associated with epiphytic plants.

The ericoid mycorrhiza is a mutualistic relationship formed between members of the plant family Ericaceae and several lineages of mycorrhizal fungi. This symbiosis represents an important adaptation to acidic and nutrient poor soils that species in the Ericaceae typically inhabit, including boreal forests, bogs, and heathlands. Molecular clock estimates suggest that the symbiosis originated approximately 140 million years ago.
Microbial inoculants also known as soil inoculants or bioinoculants are agricultural amendments that use beneficial rhizosphericic or endophytic microbes to promote plant health. Many of the microbes involved form symbiotic relationships with the target crops where both parties benefit (mutualism). While microbial inoculants are applied to improve plant nutrition, they can also be used to promote plant growth by stimulating plant hormone production. Although bacterial and fungal inoculants are common, inoculation with archaea to promote plant growth is being increasingly studied.
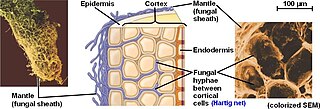
The Hartig net is the network of inward-growing hyphae, that extends into the plant host root, penetrating between plant cells in the root epidermis and cortex in ectomycorrhizal symbiosis. This network is the internal component of fungal morphology in ectomycorrhizal symbiotic structures formed with host plant roots, in addition to a hyphal mantle or sheath on the root surface, and extramatrical mycelium extending from the mantle into the surrounding soil. The Hartig net is the site of mutualistic resource exchange between the fungus and the host plant. Essential nutrients for plant growth are acquired from the soil by exploration and foraging of the extramatrical mycelium, then transported through the hyphal network across the mantle and into the Hartig net, where they are released by the fungi into the root apoplastic space for uptake by the plant. The hyphae in the Hartig net acquire sugars from the plant root, which are transported to the external mycelium to provide a carbon source to sustain fungal growth.
Nitrogen nutrition in the arbuscular mycorrhizal system refers to...
The mycorrhizosphere is the region around a mycorrhizal fungus in which nutrients released from the fungus increase the microbial population and its activities. The roots of most terrestrial plants, including most crop plants and almost all woody plants, are colonized by mycorrhiza-forming symbiotic fungi. In this relationship, the plant roots are infected by a fungus, but the rest of the fungal mycelium continues to grow through the soil, digesting and absorbing nutrients and water and sharing these with its plant host. The fungus in turn benefits by receiving photosynthetic sugars from its host. The mycorrhizosphere consists of roots, hyphae of the directly connected mycorrhizal fungi, associated microorganisms, and the soil in their direct influence.

Soil carbon storage is an important function of terrestrial ecosystems. Soil contains more carbon than plants and the atmosphere combined. Understanding what maintains the soil carbon pool is important to understand the current distribution of carbon on Earth, and how it will respond to environmental change. While much research has been done on how plants, free-living microbial decomposers, and soil minerals affect this pool of carbon, it is recently coming to light that mycorrhizal fungi—symbiotic fungi that associate with roots of almost all living plants—may play an important role in maintaining this pool as well. Measurements of plant carbon allocation to mycorrhizal fungi have been estimated to be 5 to 20% of total plant carbon uptake, and in some ecosystems the biomass of mycorrhizal fungi can be comparable to the biomass of fine roots. Recent research has shown that mycorrhizal fungi hold 50 to 70 percent of the total carbon stored in leaf litter and soil on forested islands in Sweden. Turnover of mycorrhizal biomass into the soil carbon pool is thought to be rapid and has been shown in some ecosystems to be the dominant pathway by which living carbon enters the soil carbon pool.

An ectomycorrhiza is a form of symbiotic relationship that occurs between a fungal symbiont, or mycobiont, and the roots of various plant species. The mycobiont is often from the phyla Basidiomycota and Ascomycota, and more rarely from the Zygomycota. Ectomycorrhizas form on the roots of around 2% of plant species, usually woody plants, including species from the birch, dipterocarp, myrtle, beech, willow, pine and rose families. Research on ectomycorrhizas is increasingly important in areas such as ecosystem management and restoration, forestry and agriculture.

Ectomycorrhizal extramatrical mycelium is the collection of filamentous fungal hyphae emanating from ectomycorrhizas. It may be composed of fine, hydrophilic hypha which branches frequently to explore and exploit the soil matrix or may aggregate to form rhizomorphs; highly differentiated, hydrophobic, enduring, transport structures.

The root microbiome is the dynamic community of microorganisms associated with plant roots. Because they are rich in a variety of carbon compounds, plant roots provide unique environments for a diverse assemblage of soil microorganisms, including bacteria, fungi, and archaea. The microbial communities inside the root and in the rhizosphere are distinct from each other, and from the microbial communities of bulk soil, although there is some overlap in species composition.
The fungal loop hypothesis suggests that soil fungi in arid ecosystems connect the metabolic activity of plants and biological soil crusts which respond to different soil moisture levels. Compiling diverse evidence such as limited accumulation of soil organic matter, high phenol oxidative and proteolytic enzyme potentials due to microbial activity, and symbioses between plants and fungi, the fungal loop hypothesis suggests that carbon and nutrients are cycled in biotic pools rather than leached or effluxed to the atmosphere during and between pulses of precipitation.
Orchid mycorrhizae are endomycorrhizal fungi which develop symbiotic relationships with the roots and seeds of plants of the family Orchidaceae. Nearly all orchids are myco-heterotrophic at some point in their life cycle. Orchid mycorrhizae are critically important during orchid germination, as an orchid seed has virtually no energy reserve and obtains its carbon from the fungal symbiont.

Mycorrhiza helper bacteria (MHB) are a group of organisms that form symbiotic associations with both ectomycorrhiza and arbuscular mycorrhiza. MHBs are diverse and belong to a wide variety of bacterial phyla including both Gram-negative and Gram-positive bacteria. Some of the most common MHBs observed in studies belong to the phylas Pseudomonas and Streptomyces. MHBs have been seen to have extremely specific interactions with their fungal hosts at times, but this specificity is lost with plants. MHBs enhance mycorrhizal function, growth, nutrient uptake to the fungus and plant, improve soil conductance, aid against certain pathogens, and help promote defense mechanisms. These bacteria are naturally present in the soil, and form these complex interactions with fungi as plant root development starts to take shape. The mechanisms through which these interactions take shape are not well-understood and needs further study.
Leohumicola verrucosa is a heat-resistant, endophytic, ericoid mycorrhizal soil fungus. Its species name refers to rough, warty or spine-like ornamentations on its aleurioconidia. L. verrucosa was first described from samples of soil exposed to fire; among these it was especially abundant in regularly burned blueberry fields in eastern Canada. L. verrucosa forms mycorrhizal relationships with a wide variety and distribution of species in the Ericaceae family.

Mucoromycota is a division within the kingdom fungi. It includes a diverse group of various molds, including the common bread molds Mucor and Rhizopus. It is a sister phylum to Dikarya.
Darksidea is a genus of fungi belonging to the ascomycetes, in the order Pleosporales. It was described in 2015 by D.G. Knapp, Kovács, J.Z. Groenew. & Crous. Like many other members of the ascomycetes, they are dark septate endophytes, who colonize epidermal, intra- and inter-cellular root tissue, nearly always found upon grass species in arid or semiarid climates.
References
- 1 2 3 4 Jumpponen, A. 2001. Dark-Septate Endophytes – are they mycorrhizal? Mycorrhiza 11:207–211.
- 1 2 3 4 5 6 7 8 Mandyam, K. and Jumpponen, A. 2005. Seeking the elusive function of the root-colonising dark septate endophytic fungi. Studies in Mycology. 53: 173–189.
- 1 2 3 4 5 6 7 Jumpponen, Ari; Trappe, James M. (1998). "Dark septate endophytes: a review of facultative biotrophic root-colonizing fungi". New Phytologist. 140 (2): 295–310. doi: 10.1046/j.1469-8137.1998.00265.x . ISSN 0028-646X. PMID 33862835.
- ↑ Caldwell, B.A., Jumpponen, A., Trappe, J.M. 2000. Utilization of Major Detrital Substrates by Dark-Septate, Root Endophytes. Mycologia 92: 230–232.
- ↑ Reininger, V. and Sieber, T.N. 2013. Mitigation of antagonistic effects on plant growth due to root co-colonization by dark septate endophytes and ectomycorrhiza. Environmental Microbiology Reports 5(6), 892–898
- 1 2 Newsham, K.K. 2011. A meta-analysis of plant responses to dark septate root endophytes. New Phytologist 190: 783–793.
- ↑ Porras-Alfaro, A., Herrera, J., Natvig, D.O., Lipinski, K., Sinsabaugh, R.L. 2011. Diversity and distribution of soil fungal communities in a semiarid grassland. Mycologia 103: 10–21.