This gene encodes a member of the galectin family. Galectins are beta-galactoside-binding animal lectins with conserved carbohydrate recognition domains. The galectins have been implicated in many essential functions including development, differentiation, cell-cell adhesion, cell-matrix interaction, growth regulation, apoptosis, and RNA splicing. This gene is widely expressed in tumoral tissues and seems to be involved in integrin-like cell interactions. Alternatively spliced transcript variants encoding different isoforms have been identified.[7]
Galectin-8, interacts with the mTOR regulatory system composed of SLC38A9, Ragulator, RagAB, RagCD.[8] Galectin-8 controls mTOR causing its inactivation and dissociation from damaged lysosomes, hence transducing the breach of the lysosomal membrane to mTOR.[8] The physiological consequences of mTOR inhibition following lysosomal membrane damage[8] encompass autophagy and metabolic switching.
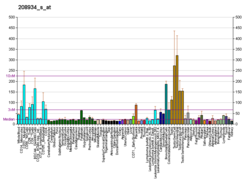
Galectin-8 levels
Circulating galectin levels in the serum of healthy individuals typically range between 0.1 and 166.3 ng/mL, with a median concentration of approximately 6 ng/mL. In cancer patients, these levels can increase up to fivefold.[9][10]
Role in cancer
Galectin-8 has a complex role in cancer, sometimes being protumorigenic and other times anti-tumorigenic.[11]
Role in cellular defence
Galectin-8 has recently been shown to have a role in cellular defence, against both bacterial cytosolic infection and vacuolar damage.[12] Many intracellular bacteria, such as S.enterica serovar Typhimurium and S.flexneri prefer to replicate inside and outside of the vacuole safety respectively, yet these vacuoles may become damaged, exposing bacteria to the host cell cytoplasm. It has been shown that the binding of galectin-8 to the damaged vacuole can recruit autophagy adaptors such as NDP52 leading to the formation of an autophagosome and subsequent bacterial destruction.[12] As knockout experiments of galectin-8 leads to more successful cytosolic replication by S.enterica serovar Typhimurium, it is thought that galectin-8 acts as a danger receptor in defence against intracellular pathogens.[12][13]
Engineered galectin-8 assays
Galectin-8 has also been used to study endosomal disruption in the development of nanoscale drug delivery systems. Many drug delivery systems carrying large molecule drugs, such as antisense oligonucleotides, siRNA, peptides, and therapeutic proteins, are engineered to be pH-responsive, and disrupt the endosomal membrane because of the lower pH found within progressively acidifying endosomes. Galectin-8 can be tagged with a fluorophore to track these disrupted endosomal membranes, especially when coupled with automated microscopy.[14]
↑ Barrow H, Guo X, Wandall HH, Pedersen JW, Fu B, Zhao Q, etal. (November 2011). "Serum galectin-2, -4, and -8 are greatly increased in colon and breast cancer patients and promote cancer cell adhesion to blood vascular endothelium". Clinical Cancer Research. 17 (22): 7035–7046. doi:10.1158/1078-0432.CCR-11-1462. PMID21933892.
1 2 3 Hadari YR, Arbel-Goren R, Levy Y, Amsterdam A, Alon R, Zakut R, Zick Y (July 2000). "Galectin-8 binding to integrins inhibits cell adhesion and induces apoptosis". Journal of Cell Science. 113 (Pt 13): 2385–2397. doi:10.1242/jcs.113.13.2385. PMID10852818.
Further reading
Bidon N, Brichory F, Bourguet P, Le Pennec JP, Dazord L (September 2001). "Galectin-8: a complex sub-family of galectins (Review)". International Journal of Molecular Medicine. 8 (3): 245–50. doi:10.3892/ijmm.8.3.245. PMID11494049.
Danguy A, Camby I, Kiss R (September 2002). "Galectins and cancer". Biochimica et Biophysica Acta (BBA) - General Subjects. 1572 (2–3): 285–93. doi:10.1016/S0304-4165(02)00315-X. PMID12223276.
Bassen R, Brichory F, Caulet-Maugendre S, Bidon N, Delaval P, Desrues B, Dazord L (2000). "Expression of Po66-CBP, a type-8 galectin, in different healthy, tumoral and peritumoral tissues". Anticancer Research. 19 (6B): 5429–33. PMID10697573.
Hadari YR, Arbel-Goren R, Levy Y, Amsterdam A, Alon R, Zakut R, Zick Y (July 2000). "Galectin-8 binding to integrins inhibits cell adhesion and induces apoptosis". Journal of Cell Science. 113 ( Pt 13) (13): 2385–97. doi:10.1242/jcs.113.13.2385. PMID10852818.
Bidon N, Brichory F, Hanash S, Bourguet P, Dazord L, Le Pennec JP (August 2001). "Two messenger RNAs and five isoforms for Po66-CBP, a galectin-8 homolog in a human lung carcinoma cell line". Gene. 274 (1–2): 253–62. doi:10.1016/S0378-1119(01)00598-4. PMID11675018.
Maier C, Rösch K, Herkommer K, Bochum S, Cancel-Tassin G, Cussenot O, etal. (September 2002). "A candidate gene approach within the susceptibility region PCaP on 1q42.2-43 excludes deleterious mutations of the PCTA-1 gene to be responsible for hereditary prostate cancer". European Urology. 42 (3): 301–7. doi:10.1016/S0302-2838(02)00280-4. PMID12234517.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.