Related Research Articles

Fluorescence is the emission of light by a substance that has absorbed light or other electromagnetic radiation. It is a form of luminescence. In most cases, the emitted light has a longer wavelength, and therefore a lower photon energy, than the absorbed radiation. A perceptible example of fluorescence occurs when the absorbed radiation is in the ultraviolet region of the electromagnetic spectrum, while the emitted light is in the visible region; this gives the fluorescent substance a distinct color that can only be seen when the substance has been exposed to UV light. Fluorescent materials cease to glow nearly immediately when the radiation source stops, unlike phosphorescent materials, which continue to emit light for some time after.
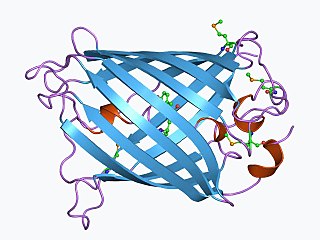
The green fluorescent protein (GFP) is a protein that exhibits green fluorescence when exposed to light in the blue to ultraviolet range. The label GFP traditionally refers to the protein first isolated from the jellyfish Aequorea victoria and is sometimes called avGFP. However, GFPs have been found in other organisms including corals, sea anemones, zoanithids, copepods and lancelets.

In molecular biology and biotechnology, a fluorescent tag, also known as a fluorescent label or fluorescent probe, is a molecule that is attached chemically to aid in the detection of a biomolecule such as a protein, antibody, or amino acid. Generally, fluorescent tagging, or labeling, uses a reactive derivative of a fluorescent molecule known as a fluorophore. The fluorophore selectively binds to a specific region or functional group on the target molecule and can be attached chemically or biologically. Various labeling techniques such as enzymatic labeling, protein labeling, and genetic labeling are widely utilized. Ethidium bromide, fluorescein and green fluorescent protein are common tags. The most commonly labelled molecules are antibodies, proteins, amino acids and peptides which are then used as specific probes for detection of a particular target.

Fluorescence spectroscopy is a type of electromagnetic spectroscopy that analyzes fluorescence from a sample. It involves using a beam of light, usually ultraviolet light, that excites the electrons in molecules of certain compounds and causes them to emit light; typically, but not necessarily, visible light. A complementary technique is absorption spectroscopy. In the special case of single molecule fluorescence spectroscopy, intensity fluctuations from the emitted light are measured from either single fluorophores, or pairs of fluorophores.
A fluorophore is a fluorescent chemical compound that can re-emit light upon light excitation. Fluorophores typically contain several combined aromatic groups, or planar or cyclic molecules with several π bonds.

Fura-2, an aminopolycarboxylic acid, is a ratiometric fluorescent dye which binds to free intracellular calcium. It was the first widely used dye for calcium imaging, and remains very popular. Fura-2 is excited at 340 nm and 380 nm of light, and the ratio of the emissions at those wavelengths is directly related to the amount of intracellular calcium. Regardless of the presence of calcium, Fura-2 emits at 510 nm of light. The use of the ratio automatically cancels out confounding variables, such as variable dye concentration and cell thickness, making Fura-2 one of the most appreciated tools to quantify calcium levels. The high photon yield of fura-2 allowed the first real time measurements of calcium inside living cells in 1986. More recently, genetically encoded calcium indicators based on spectral variants of the green fluorescent protein, such as Cameleons, have supplemented the use of Fura-2 and other small molecule dyes for calcium imaging, but Fura-2 remains faster.

A fluorescence microscope is an optical microscope that uses fluorescence instead of, or in addition to, scattering, reflection, and attenuation or absorption, to study the properties of organic or inorganic substances. "Fluorescence microscope" refers to any microscope that uses fluorescence to generate an image, whether it is a simple set up like an epifluorescence microscope or a more complicated design such as a confocal microscope, which uses optical sectioning to get better resolution of the fluorescence image.

Texas Red or sulforhodamine 101 acid chloride is a red fluorescent dye, used in histology for staining cell specimens, for sorting cells with fluorescent-activated cell sorting machines, in fluorescence microscopy applications, and in immunohistochemistry. Texas Red fluoresces at about 615 nm, and the peak of its absorption spectrum is at 589 nm. The powder is dark purple. Solutions can be excited by a dye laser tuned to 595-605 nm, or less efficiently a krypton laser at 567 nm. The absorption extinction coefficient at 596 nm is about 85,000 M−1cm−1.
EosFP is a photoactivatable green to red fluorescent protein. Its green fluorescence (516 nm) switches to red (581 nm) upon UV irradiation of ~390 nm due to a photo-induced modification resulting from a break in the peptide backbone near the chromophore. Eos was first discovered as a tetrameric protein in the stony coral Lobophyllia hemprichii. Like other fluorescent proteins, Eos allows for applications such as the tracking of fusion proteins, multicolour labelling and tracking of cell movement. Several variants of Eos have been engineered for use in specific study systems including mEos2, mEos4 and CaMPARI.
Photoactivatable fluorescent proteins (PAFPs) is a type of fluorescent protein that exhibit fluorescence that can be modified by a light-induced chemical reaction.

Fluorescence is used in the life sciences generally as a non-destructive way of tracking or analysing biological molecules. Some proteins or small molecules in cells are naturally fluorescent, which is called intrinsic fluorescence or autofluorescence. Alternatively, specific or general proteins, nucleic acids, lipids or small molecules can be "labelled" with an extrinsic fluorophore, a fluorescent dye which can be a small molecule, protein or quantum dot. Several techniques exist to exploit additional properties of fluorophores, such as fluorescence resonance energy transfer, where the energy is passed non-radiatively to a particular neighbouring dye, allowing proximity or protein activation to be detected; another is the change in properties, such as intensity, of certain dyes depending on their environment allowing their use in structural studies.

GCaMP is a genetically encoded calcium indicator (GECI) initially developed in 2001 by Junichi Nakai. It is a synthetic fusion of green fluorescent protein (GFP), calmodulin (CaM), and M13, a peptide sequence from myosin light-chain kinase. When bound to Ca2+, GCaMP fluoresces green with a peak excitation wavelength of 480 nm and a peak emission wavelength of 510 nm. It is used in biological research to measure intracellular Ca2+ levels both in vitro and in vivo using virally transfected or transgenic cell and animal lines. The genetic sequence encoding GCaMP can be inserted under the control of promoters exclusive to certain cell types, allowing for cell-type specific expression of GCaMP. Since Ca2+ is a second messenger that contributes to many cellular mechanisms and signaling pathways, GCaMP allows researchers to quantify the activity of Ca2+-based mechanisms and study the role of Ca2+ ions in biological processes of interest.
mCherry is a member of the mFruits family of monomeric red fluorescent proteins (mRFPs). As a RFP, mCherry was derived from DsRed of Discosoma sea anemones unlike green fluorescent proteins (GFPs) which are often derived from Aequorea victoria jellyfish. Fluorescent proteins are used to tag components in the cell, so they can be studied using fluorescence spectroscopy and fluorescence microscopy. mCherry absorbs light between 540-590 nm and emits light in the range of 550-650 nm. mCherry belongs to the group of fluorescent protein chromophores used as instruments to visualize genes and analyze their functions in experiments. Genome editing has been improved greatly through the precise insertion of these fluorescent protein tags into the genetic material of many diverse organisms. Most comparisons between the brightness and photostability of different fluorescent proteins have been made in vitro, removed from biological variables that affect protein performance in cells or organisms. It is hard to perfectly simulate cellular environments in vitro, and the difference in environment could have an effect on the brightness and photostability.
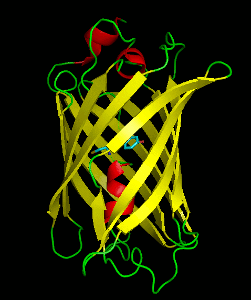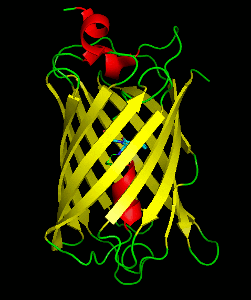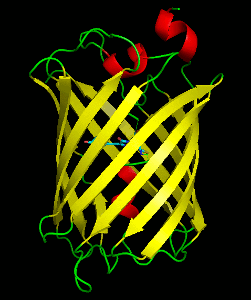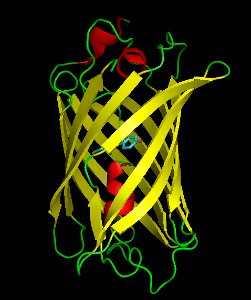
Dronpa is a reversibly switchable photoactivatable fluorescent protein that is 2.5 times as bright as EGFP. Dronpa gets switched off by strong illumination with 488 nm (blue) light and this can be reversed by weak 405 nm UV light. A single dronpa molecule can be switched on and off over 100 times. It has an excitation peak at 503 nm and an emission peak at 518 nm.

Photoactivatable probes, or caged probes, are cellular players that can be triggered by a flash of light. They are used in biological research to study processes in cells. The basic principle is to bring a photoactivatable agent to cells, tissues or even living animals and specifically control its activity by illumination.
Calcium imaging is a microscopy technique to optically measure the calcium (Ca2+) status of an isolated cell, tissue or medium. Calcium imaging takes advantage of calcium indicators, fluorescent molecules that respond to the binding of Ca2+ ions by fluorescence properties. Two main classes of calcium indicators exist: chemical indicators and genetically encoded calcium indicators (GECI). This technique has allowed studies of calcium signalling in a wide variety of cell types. In neurons, action potential generation is always accompanied by rapid influx of Ca2+ ions. Thus, calcium imaging can be used to monitor the electrical activity in hundreds of neurons in cell culture or in living animals, which has made it possible to observe the activity of neuronal circuits during ongoing behavior.

A FMN-binding fluorescent protein (FbFP), also known as a LOV-based fluorescent protein, is a small, oxygen-independent fluorescent protein that binds flavin mononucleotide (FMN) as a chromophore.

Small ultra red fluorescent protein (smURFP) is a class of far-red fluorescent protein evolved from a cyanobacterial phycobiliprotein, α-allophycocyanin. Native α-allophycocyanin requires an exogenous protein, known as a lyase, to attach the chromophore, phycocyanobilin. Phycocyanobilin is not present in mammalian cells. smURFP was evolved to covalently attach phycocyanobilin without a lyase and fluoresce, covalently attach biliverdin and fluoresce, blue-shift fluorescence to match the organic fluorophore, Cy5, and not inhibit E. coli growth. smURFP was found after 12 rounds of random mutagenesis and manually screening 10,000,000 bacterial colonies.

Red fluorescent protein (RFP) is a fluorophore that fluoresces red-orange when excited. Several variants have been developed using directed mutagenesis. The original was isolated from Discosoma, and named DsRed. Others are now available that fluoresce orange, red, and far-red.
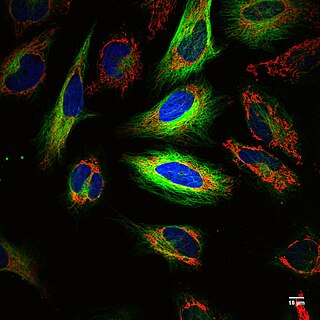
Fluorescence imaging is a type of non-invasive imaging technique that can help visualize biological processes taking place in a living organism. Images can be produced from a variety of methods including: microscopy, imaging probes, and spectroscopy.
References
- Tomura, M.; Yoshida, N.; Tanaka, J.; Karasawa, S.; Miwa, Y.; Miyawaki, A.; Kanagawa, O. (2008). "Monitoring cellular movement in vivo with photoconvertible fluorescence protein "Kaede" transgenic mice". Proceedings of the National Academy of Sciences. 105 (31): 10871–10876. Bibcode:2008PNAS..10510871T. doi: 10.1073/pnas.0802278105 . PMC 2504797 . PMID 18663225.
- Dittrich, P. S.; Schäfer, S. P.; Schwille, P. (2005). "Characterization of the Photoconversion on Reaction of the Fluorescent Protein Kaede on the Single-Molecule Level". Biophysical Journal. 89 (5): 3446–3455. Bibcode:2005BpJ....89.3446D. doi:10.1529/biophysj.105.061713. PMC 1366840 . PMID 16055537.
- 1 2 3 Ando, R. (2002). "An optical marker based on the UV-induced green-to-red photoconversion of a fluorescent protein". Proceedings of the National Academy of Sciences. 99 (20): 12651–12656. Bibcode:2002PNAS...9912651A. doi: 10.1073/pnas.202320599 . PMC 130515 . PMID 12271129.
- ↑ Mizuno, H.; Mal, T. K.; Tong, K. I.; Ando, R.; Furuta, T.; Ikura, M.; Miyawaki, A. (2003). "Photo-induced peptide cleavage in the green-to-red conversion of a fluorescent protein". Molecular Cell. 12 (4): 1051–1058. doi: 10.1016/s1097-2765(03)00393-9 . PMID 14580354.
- 1 2 Lippincott-Schwartz, J.; Altan-Bonnet, N.; Patterson, G. H. (September 2003). "Photobleaching and photoactivation: Following protein dynamics in living cells". Nature Cell Biology. Suppl: S7–14. CiteSeerX 10.1.1.1072.3400 . doi:10.1038/ncb1032 (inactive 31 January 2024). PMID 14562845. S2CID 15044107.
{{cite journal}}: CS1 maint: DOI inactive as of January 2024 (link) CS1 maint: date and year (link) - ↑ Mutoh, T.; Miyata, T.; Kashiwagi, S.; Miyawaki, A.; Ogawa, M. (2006). "Dynamic behavior of individual cells in developing organotypic brain slices revealed by the photoconvertable protein Kaede". Experimental Neurology. 200 (2): 430–437. doi:10.1016/j.expneurol.2006.03.022. PMID 16753144. S2CID 22238647.