Related Research Articles

The molars or molar teeth are large, flat teeth at the back of the mouth. They are more developed in mammals. They are used primarily to grind food during chewing. The name molar derives from Latin, molaris dens, meaning "millstone tooth", from mola, millstone and dens, tooth. Molars show a great deal of diversity in size and shape across the mammal groups. The third molar of humans is sometimes vestigial.

Gondwanatheria is an extinct group of mammaliaforms that lived in parts of Gondwana, including Madagascar, India, South America, Africa, and Antarctica during the Upper Cretaceous through the Miocene. Until recently, they were known only from fragmentary remains. They are generally considered to be closely related to the multituberculates and likely the euharamiyidians, well known from the Northern Hemisphere, with which they form the clade Allotheria.
Ferugliotherium is a genus of fossil mammals in the family Ferugliotheriidae from the Campanian and/or Maastrichtian period of Argentina. It contains a single species, Ferugliotherium windhauseni, which was first described in 1986. Although originally interpreted on the basis of a single brachydont (low-crowned) molar as a member of Multituberculata, an extinct group of small, rodent-like mammals, it was recognized as related to the hypsodont (high-crowned) Sudamericidae following the discovery of additional material in the early 1990s. After a jaw of the sudamericid Sudamerica was described in 1999, these animals were no longer considered to be multituberculates and a few fossils that were previously considered to be Ferugliotherium were assigned to unspecified multituberculates instead. Since 2005, a relationship between gondwanatheres and multituberculates has again received support. A closely related animal, Trapalcotherium, was described in 2009 on the basis of a single tooth.
Lavanify is a mammalian genus from the late Cretaceous of Madagascar. The only species, L. miolaka, is known from two isolated teeth, one of which is damaged. The teeth were collected in 1995–1996 and described in 1997. The animal is classified as a member of Gondwanatheria, an enigmatic extinct group with unclear phylogenetic relationships, and within Gondwanatheria as a member of the family Sudamericidae. Lavanify is most closely related to the Indian Bharattherium; the South American Sudamerica and Gondwanatherium are more distantly related. Gondwanatheres probably ate hard plant material.
Ferugliotheriidae is one of three known families in the order Gondwanatheria, an enigmatic group of extinct mammals. Gondwanatheres have been classified as a group of uncertain affinities or as members of Multituberculata, a major extinct mammalian order. The best-known representative of Ferugliotheriidae is the genus Ferugliotherium from the Late Cretaceous epoch in Argentina. A second genus, Trapalcotherium, is known from a single tooth, a first lower molariform, from a different Late Cretaceous Argentinean locality. Another genus known from a single tooth, Argentodites, was first described as an unrelated multituberculate, but later identified as possibly related to Ferugliotherium. Finally, a single tooth from the Paleogene of Peru, LACM 149371, perhaps a last upper molariform, and a recent specimen from Mexico, may represent related animals.

Ambondro mahabo is a mammal from the Middle Jurassic (Bathonian) Isalo III Formation of Madagascar. The only described species of the genus Ambondro, it is known from a fragmentary lower jaw with three teeth, interpreted as the last premolar and the first two molars. The premolar consists of a central cusp with one or two smaller cusps and a cingulum (shelf) on the inner, or lingual, side of the tooth. The molars also have such a lingual cingulum. They consist of two groups of cusps: a trigonid of three cusps at the front and a talonid with a main cusp, a smaller cusp, and a crest at the back. Features of the talonid suggest that Ambondro had tribosphenic molars, the basic arrangement of molar features also present in marsupial and placental mammals. It is the oldest known mammal with putatively tribosphenic teeth; at the time of its discovery it antedated the second oldest example by about 25 million years.
Many different terms have been proposed for features of the tooth crown in mammals.
UA 8699 is a fossil mammalian tooth from the Cretaceous of Madagascar. A broken lower molar about 3.5 mm (0.14 in) long, it is from the Maastrichtian of the Maevarano Formation in northwestern Madagascar. Details of its crown morphology indicate that it is a boreosphenidan, a member of the group that includes living marsupials and placental mammals. David W. Krause, who first described the tooth in 2001, interpreted it as a marsupial on the basis of five shared characters, but in 2003 Averianov and others noted that all those are shared by zhelestid placentals and favored a close relationship between UA 8699 and the Spanish zhelestid Lainodon. Krause used the tooth as evidence that marsupials were present on the southern continents (Gondwana) as early as the late Cretaceous and Averianov and colleagues proposed that the tooth represented another example of faunal exchange between Africa and Europe at the time.

Brachytarsomys mahajambaensis is an extinct rodent from northwestern Madagascar. It is known from nine isolated molars found in several sites during fieldwork that started in 2001. First described in 2010, it is placed in the genus Brachytarsomys together with two larger living species, which may differ in some details of molar morphology. The presence of B. mahajambaensis, a rare element in the local rodent fauna, suggests that the region was previously more humid.

Nesomys narindaensis is an extinct rodent that lived in northwestern Madagascar. It is known from subfossil skull bones and isolated molars found in several sites during field work that started in 2001. First described in 2010, it is placed in the genus Nesomys together with three smaller living species, which may differ in some details of molar morphology. The presence of N. narindaensis, a rare element in the local rodent fauna, suggests that the region was previously more humid.
Several mammals are known from the Mesozoic of Madagascar. The Bathonian Ambondro, known from a piece of jaw with three teeth, is the earliest known mammal with molars showing the modern, tribosphenic pattern that is characteristic of marsupial and placental mammals. Interpretations of its affinities have differed; one proposal places it in a group known as Australosphenida with other Mesozoic tribosphenic mammals from the southern continents (Gondwana) as well as the monotremes, while others favor closer affinities with northern (Laurasian) tribosphenic mammals or specifically with placentals. At least five species are known from the Maastrichtian, including a yet undescribed species known from a nearly complete skeleton that may represent a completely new group of mammals. The gondwanathere Lavanify, known from two teeth, is most closely related to other gondwanatheres found in India and Argentina. Two other teeth may represent another gondwanathere or a different kind of mammal. One molar fragment is one of the few known remains of a multituberculate mammal from Gondwana and another has been interpreted as either a marsupial or a placental.
Argentodites is a possible multituberculate mammal from the Cretaceous of Argentina. The single species, Argentodites coloniensis, is known from a single blade-like fourth lower premolar (p4) from the La Colonia Formation, which is mostly or entirely Maastrichtian in age. The p4 is 4.15 mm long and bears eight cusps on its upper margin and long associated ridges on both sides. The enamel consists of prisms that are completely or partly surrounded by a sheath and that are on average 6.57 μm apart. Zofia Kielan-Jaworowska, who described and named the fossil in 2007, regarded it as a multituberculate, perhaps a cimolodontan—and thus, a member of a mostly Laurasian (northern) group and an immigrant to Argentina from North America—on the basis of the shape of the tooth and features of its enamel. In 2009, however, two teams argued that Argentodites may in fact be close to or identical with Ferugliotherium, a member of the small Gondwanan (southern) group Gondwanatheria; although their relationships are disputed, gondwanatheres may themselves be multituberculates.
Trapalcotherium is a fossil mammal from the Cretaceous of Argentina in the family Ferugliotheriidae. The single species, T. matuastensis, is known from one tooth, a first lower molar. It is from the Allen Formation, which is probably Maastrichtian in age, and was first described in 2009. The tooth bears two rows of cusps, one at the inner (lingual) side and the other at the outer (labial) side, which are connected by transverse ridges separated by deep valleys. This pattern is reminiscent of Ferugliotherium, a gondwanathere mammal from similarly aged deposits in Argentina, and Trapalcotherium is therefore recognized as a member of the same family Ferugliotheriidae. Ferugliotheriidae is one of two families of gondwanatheres, an enigmatic group without close relationships to any living mammals.
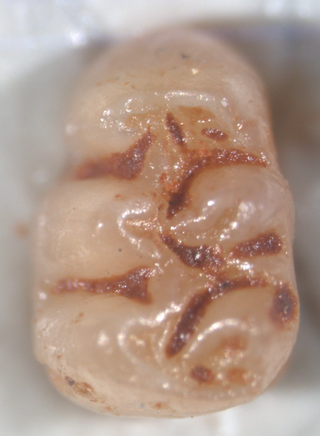
Agathaeromys is an extinct genus of oryzomyine rodents from the Pleistocene of Bonaire, Netherlands Antilles. Two species are known, which differ in size and some details of tooth morphology. The larger A. donovani, the type species, is known from hundreds of teeth that are probably 900,000 to 540,000 years old, found in four localities. A. praeuniversitatis, the smaller species, is known from 35 teeth found in a single fossil site, which is probably 540,000 to 230,000 years old.
Bharattherium is a mammal that lived in India during the Maastrichtian and possibly the Paleocene. The genus has a single species, Bharattherium bonapartei. It is part of the gondwanathere family Sudamericidae, which is also found in Madagascar and South America during the latest Cretaceous. The first fossil of Bharattherium was discovered in 1989 and published in 1997, but the animal was not named until 2007, when two teams independently named the animal Bharattherium bonapartei and Dakshina jederi. The latter name is now a synonym. Bharattherium is known from a total of eight isolated fossil teeth, including one incisor and seven molariforms.
? Nycticebus linglom is a fossil strepsirrhine primate from the Miocene of Thailand. Known only from a single tooth, an upper third molar, it is thought to be related to the living slow lorises, but the material is not sufficient to assign the species to Nycticebus with certainty, and the species name therefore uses open nomenclature. With a width of 1.82 mm, this tooth is very small for a primate. It is triangular in shape, supported by a single root, and shows three main cusps, in addition to various crests. The absence of a fourth cusp, the hypocone, distinguishes it from various other prosimian primates.
Tupaia miocenica is a fossil treeshrew from the Miocene of Thailand. Known only from a single tooth, an upper first or second molar, it is among the few known fossil treeshrews. With a length of 3.57 mm, the tooth is large for a treeshrew. At the back lingual corner, the tooth shows a small cusp, the hypocone, that is separated from the protocone in front of it by a narrow valley. The condition of the hypocone distinguishes this species from various other treeshrews. In addition, the presence of a well-developed but simple mesostyle is distinctive.
Dermotherium is a genus of fossil mammals closely related to the living colugos, a small group of gliding mammals from Southeast Asia. Two species are recognized: D. major from the Late Eocene of Thailand, based on a single fragment of the lower jaw, and D. chimaera from the Late Oligocene of Thailand, known from three fragments of the lower jaw and two isolated upper molars. In addition, a single isolated upper molar from the Early Oligocene of Pakistan has been tentatively assigned to D. chimaera. All sites where fossils of Dermotherium have been found were probably forested environments and the fossil species were probably forest dwellers like living colugos, but whether they had the gliding adaptations of the living species is unknown.
The Santa Rosa local fauna consists of the animals found in the Paleogene fossil site of Santa Rosa in eastern Peru. The age of the Santa Rosa fauna is difficult to determine, but may be Eocene (Mustersan) or Oligocene (Deseadan).
Groeberiidae is a family of strange non-placental mammals from the Eocene and Oligocene epochs of Patagonia, Argentina and Chile, South America. Originally classified as paucituberculate marsupials, they were suggested to be late representatives of the allothere clade Gondwanatheria. However, the relationship of the type genus, Groeberia, to Gondwanatheria has been firmly rejected by other scholars.
References
- 1 2 3 Goin et al., 2004, p. 145
- ↑ Campbell, 2004, pp. 156–159
- ↑ Goin et al., 2004
- ↑ Campbell, 2004, pp. 159–160
- ↑ Vucetich et al., 2010, pp. 201–202
- ↑ Campbell, 2004, p. 161
- 1 2 Goin et al., 2004, p. 146
- 1 2 3 4 5 6 Goin et al., 2004, p. 149
- ↑ Goin et al., 2004, p. 146, fig. 2
- ↑ Goin et al., 2004, p. 147
- ↑ Goin et al., 2004, pp. 147–148
- ↑ Goin et al., 2004, pp. 148–149
- ↑ Goin et al., 2004, p. 148
- 1 2 Goin et al., 2004, p. 151
- ↑ Goin et al., 2004, pp. 151–152
- 1 2 Goin et al., 2004, p. 152
Bibliography
- Campbell, K.E., Jr. 2004. The Santa Rosa local fauna: A summary. Science Series, Natural History Museum of Los Angeles County 40:155–163.
- Goin, F.J., Vieytes, E.C., Vucetich, M.G., Carlini, A.A. and Bond, M. 2004. Enigmatic mammal from the Paleogene of Perú. Science Series, Natural History Museum of Los Angeles County 40:145–153.
- Vucetich, M.G., Vieytes, E.C., Pérez, M.E. and Carloni, A.A. 2010. The rodents from La Cantera and the early evolution of caviomorphs in South America. pp. 193–205 in Madden, R.H., Carlini, A.A., Vucetich, M.G. and Kay, R.F. (eds.). The Paleontology of Gran Barranca: Evolution and Environmental Change Through the Middle Cenozoic of Patagonia. Cambridge University Press, 458 pp. ISBN 978-0-521-87241-6
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||

