
Tyrannosaurus is a genus of large theropod dinosaur. The type species Tyrannosaurus rex, often shortened to T. rex or colloquially T-Rex, is one of the best represented theropods. It lived throughout what is now western North America, on what was then an island continent known as Laramidia. Tyrannosaurus had a much wider range than other tyrannosaurids. Fossils are found in a variety of rock formations dating to the latest Campanian-Maastrichtian ages of the late Cretaceous period, 72.7 to 66 million years ago. It was the last known member of the tyrannosaurids and among the last non-avian dinosaurs to exist before the Cretaceous–Paleogene extinction event.

Tyrannosauridae is a family of coelurosaurian theropod dinosaurs that comprises two subfamilies containing up to fifteen genera, including the eponymous Tyrannosaurus. The exact number of genera is controversial, with some experts recognizing as few as three. All of these animals lived near the end of the Cretaceous Period and their fossils have been found only in North America and Asia.

Theropoda whose members are known as theropods, is an extant dinosaur clade that is characterized by hollow bones and three toes and claws on each limb. Theropods are generally classed as a group of saurischian dinosaurs. They were ancestrally carnivorous, although a number of theropod groups evolved to become herbivores and omnivores. Theropods first appeared during the Carnian age of the late Triassic period 231.4 million years ago (Ma) and included the majority of large terrestrial carnivores from the Early Jurassic until at least the close of the Cretaceous, about 66 Ma. In the Jurassic, birds evolved from small specialized coelurosaurian theropods, and are today represented by about 11,000 living species.

Spinosaurus is a genus of spinosaurid dinosaur that lived in what now is North Africa during the Cenomanian stage of the Late Cretaceous period, about 100 to 94 million years ago. The genus was known first from Egyptian remains discovered in 1912 and described by German palaeontologist Ernst Stromer in 1915. The original remains were destroyed in World War II, but additional material came to light in the early 21st century. It is unclear whether one or two species are represented in the fossils reported in the scientific literature. The best known species is S. aegyptiacus from Egypt, although a potential second species, S. maroccanus, has been recovered from Morocco. The contemporary spinosaurid genus Sigilmassasaurus has also been synonymized by some authors with S. aegyptiacus, though other researchers propose it to be a distinct taxon. Another possible junior synonym is Oxalaia from the Alcântara Formation in Brazil.

Irritator is a genus of spinosaurid dinosaur that lived in what is now Brazil during the Albian stage of the Early Cretaceous Period, about 113 to 110 million years ago. It is known from a nearly complete skull found in the Romualdo Formation of the Araripe Basin. Fossil dealers had acquired this skull and sold it to the State Museum of Natural History Stuttgart. In 1996, the specimen became the holotype of the type species Irritator challengeri. The genus name comes from the word "irritation", reflecting the feelings of paleontologists who found the skull had been heavily damaged and altered by the collectors. The species name is a homage to the fictional character Professor Challenger from Arthur Conan Doyle's novels.

Carcharodontosaurus from the “Carcharodon” lizard “This lizard was named for the Greek Carcharodon, the great white shark.” Named by German paleontologist Ernst Stromer von Reichenbach in 1931, is a genus of carnivorous theropod dinosaur that lived in North Africa from about 100 to 94 million years ago during the Cenomanian age of the Late Cretaceous. Two teeth of the genus, now lost, were first described from Algeria by French paleontologists Charles Depéret and Justin Savornin as Megalosaurus saharicus. A partial skeleton was collected by crews of German paleontologist Ernst Stromer during a 1914 expedition to Egypt. Stromer did not report the Egyptian find until 1931, in which he dubbed the novel genus Carcharodontosaurus, making the type species C. saharicus. Unfortunately, this skeleton was destroyed during the Second World War. In 1995 a nearly complete skull of C. saharicus, the first well-preserved specimen to be found in almost a century, was discovered in the Kem Kem Beds of Morocco; it was designated the neotype in 1996. Fossils unearthed from the Echkar Formation of northern Niger were described and named as another species, C. iguidensis, in 2007.

Suchomimus is a genus of spinosaur dinosaur that lived between 125 and 112 million years ago in what is now Niger, North Africa, during the Aptian to early Albian stages of the Early Cretaceous period. It was named and described by paleontologist Paul Sereno and colleagues in 1998, based on a partial skeleton from the Elrhaz Formation. Suchomimus's long and shallow skull, similar to that of a crocodile, earns it its generic name, while the specific name Suchomimus tenerensis alludes to the locality of its first remains, the Ténéré Desert.

Spinosauridae is a clade or family of tetanuran theropod dinosaurs comprising ten to seventeen known genera. Spinosaurid fossils have been recovered worldwide, including Africa, Europe, South America and Asia. Their remains have generally been attributed to the Early to Mid Cretaceous.

Tyrannosauroidea is a superfamily of coelurosaurian theropod dinosaurs that includes the family Tyrannosauridae as well as more basal relatives. Tyrannosauroids lived on the Laurasian supercontinent beginning in the Jurassic Period. By the end of the Cretaceous Period, tyrannosauroids were the dominant large predators in the Northern Hemisphere, culminating in the gigantic Tyrannosaurus. Fossils of tyrannosauroids have been recovered on what are now the continents of North America, Europe and Asia, with fragmentary remains possibly attributable to tyrannosaurs also known from South America and Australia.

Cristatusaurus is a genus of theropod dinosaur that lived during the Early Cretaceous Period of what is now Niger, 112 million years ago. It was a baryonychine member of the Spinosauridae, a group of large bipedal carnivores with well-built forelimbs and elongated, crocodile-like skulls. The type species Cristatusaurus lapparenti was named in 1998 by scientists Philippe Taquet and Dale Russell, on the basis of jaw bones and some vertebrae. Two claw fossils were also later assigned to Cristatusaurus. The animal's generic name, which means "crested reptile", alludes to a sagittal crest on top of its snout; while the specific name is in honor of the French paleontologist Albert-Félix de Lapparent. Cristatusaurus is known from the Albian to Aptian Elrhaz Formation, where it would have coexisted with sauropod and iguanodontian dinosaurs, other theropods, and various crocodylomorphs.

Poposaurus is an extinct genus of pseudosuchian archosaur from the Late Triassic of the southwestern and eastern United States. It belongs to the clade Poposauroidea, an unusual group of Triassic pseudosuchians that includes sail-backed, beaked, and aquatic forms. Fossils have been found in Wyoming, Utah, Arizona, Texas, and Virginia. Except for the skull, most parts of the skeleton are known. The type species, P. gracilis, was described and named by Maurice Goldsmith Mehl in 1915. A second species, P. langstoni, was originally the type species of the genus Lythrosuchus. Since it was first described, Poposaurus has been variously classified as a dinosaur, a phytosaur, and a "rauisuchian".

The scientific question of within which larger group of animals birds evolved has traditionally been called the "origin of birds". The present scientific consensus is that birds are a group of maniraptoran theropod dinosaurs that originated during the Mesozoic Era.
Lawrence M. Witmer is an American paleontologist and paleobiologist. He is a Professor of Anatomy and a Chang Ying-Chien Professor of Paleontology at the Department of Biomedical Sciences at the Heritage College of Osteopathic Medicine in Ohio University. Witmer is considered to be one of the world's foremost experts on soft-tissue anatomy of the skull in extinct animals such as dinosaurs and pterosaurs. One of Witmer's largest contributions to vertebrate paleontology was the creation of the Extant Phylogenetic Bracket approach to ancestral character state reconstruction. This methodology incorporates extensive comparative anatomy using physical and digital dissections of extant animals.
Reptiles arose about 320 million years ago during the Carboniferous period. Reptiles, in the traditional sense of the term, are defined as animals that have scales or scutes, lay land-based hard-shelled eggs, and possess ectothermic metabolisms. So defined, the group is paraphyletic, excluding endothermic animals like birds that are descended from early traditionally-defined reptiles. A definition in accordance with phylogenetic nomenclature, which rejects paraphyletic groups, includes birds while excluding mammals and their synapsid ancestors. So defined, Reptilia is identical to Sauropsida.

Oxalaia is a genus of spinosaurid dinosaur that lived in what is now the Northeast Region of Brazil during the Cenomanian stage of the Late Cretaceous period, sometime between 100.5 and 93.9 million years ago. Its only known fossils were found in 1999 on Cajual Island in the rocks of the Alcântara Formation, which is known for its abundance of fragmentary, isolated fossil specimens. The remains of Oxalaia were described in 2011 by Brazilian palaeontologist Alexander Kellner and colleagues, who assigned the specimens to a new genus containing one species, Oxalaia quilombensis. The species name refers to the Brazilian quilombo settlements. Oxalaia quilombensis is the eighth officially named theropod species from Brazil and the largest carnivorous dinosaur discovered there. It is closely related to the African genus Spinosaurus, and/or may be a junior synonym of this taxon.

Vegaviidae is an extinct family of birds of uncertain phylogenetic placement, which existed during the Late Cretaceous and possibly the Paleocene. Definitive fossils attributed to the family have only been found in Antarctica, though other fossils from the Southern Hemisphere including Chile and New Zealand may represent this group. The putative Campanian vegaviid from Canada known as Maaqwi, more likely belongs to the Procellariformes.
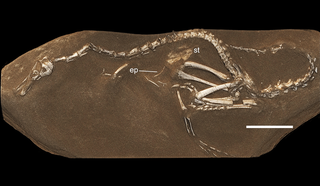
Halszkaraptor is a genus of waterfowl-like dromaeosaurid dinosaurs from Mongolia that lived during the Late Cretaceous period. It contains only one known species, Halszkaraptor escuilliei.
This article records new taxa of fossil archosaurs of every kind that are scheduled described during the year 2019, as well as other significant discoveries and events related to paleontology of archosaurs that are scheduled to occur in the year 2019.

Wulong is a genus of microraptorine dromaeosaurid dinosaurs from the Early Cretaceous (Aptian) Jiufotang Formation of China. The genus includes a single species, Wulong bohaiensis. The skeletal remains, which include preserved feathers, represent a juvenile.

Ceratosuchops is a genus of spinosaurid from the Early Cretaceous (Barremian) of Britain.
