The Balts or Baltic peoples are a group of peoples inhabiting the eastern coast of the Baltic Sea who speak Baltic languages. Among the Baltic peoples are modern-day Lithuanians and Latvians — all East Balts — as well as the Old Prussians, Curonians, Sudovians, Skalvians, Yotvingians and Galindians — the West Balts — whose languages and cultures are now extinct.

The Funnel(-neck-)beaker culture, in short TRB or TBK, was an archaeological culture in north-central Europe. It developed as a technological merger of local neolithic and mesolithic techno-complexes between the lower Elbe and middle Vistula rivers. These predecessors were the (Danubian) Lengyel-influenced Stroke-ornamented ware culture (STK) groups/Late Lengyel and Baden-Boleráz in the southeast, Rössen groups in the southwest and the Ertebølle-Ellerbek groups in the north. The TRB introduced farming and husbandry as major food sources to the pottery-using hunter-gatherers north of this line.
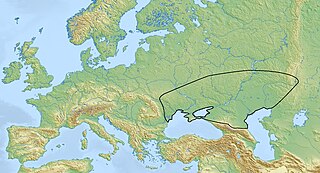
The Yamnaya culture or the Yamna culture, also known as the Pit Grave culture or Ochre Grave culture, is a late Copper Age to early Bronze Age archaeological culture of the region between the Southern Bug, Dniester, and Ural rivers, dating to 3300–2600 BCE. It was discovered by Vasily Gorodtsov following his archaeological excavations near the Donets River in 1901–1903. Its name derives from its characteristic burial tradition: Я́мная is a Russian adjective that means 'related to pits ', as these people used to bury their dead in tumuli (kurgans) containing simple pit chambers.

The Sredny Stog culture or Serednii Stih culture is a pre-Kurgan archaeological culture from the 5th–4th millennia BC. It is named after the Dnieper river islet of today's Serednii Stih, Ukraine, where it was first located.
The Pitted Ware culture was a hunter-gatherer culture in southern Scandinavia, mainly along the coasts of Svealand, Götaland, Åland, north-eastern Denmark and southern Norway. Despite its Mesolithic economy, it is by convention classed as Neolithic, since it falls within the period in which farming reached Scandinavia. The Pitted Ware people were largely maritime hunters, and were engaged in lively trade with both the agricultural communities of the Scandinavian interior and other hunter-gatherers of the Baltic Sea.
The Narva culture or eastern Baltic was a European Neolithic archaeological culture in present-day Estonia, Latvia, Lithuania, Kaliningrad Oblast, and adjacent portions of Poland, Belarus and Russia. A successor of the Mesolithic Kunda culture, the Narva culture continued up to the start of the Bronze Age. The culture spanned from c. 5300 to 1750 BC. The technology was that of hunter-gatherers. The culture was named after the Narva River in Estonia.

Cheddar Man is a human male skeleton found in Gough's Cave in Cheddar Gorge, Somerset, England. The skeletal remains date to around the mid-to-late 9th millennium BC, corresponding to the Mesolithic period, and it appears that he died a violent death. A large crater-like lesion just above the skull's right orbit suggests that the man may have also been suffering from a bone infection.
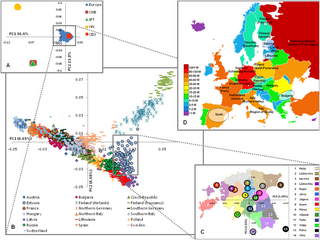
The genetic history of Europe includes information around the formation, ethnogenesis, and other DNA-specific information about populations indigenous, or living in Europe.
The Kunda culture, which originated from the Swiderian culture, comprised Mesolithic hunter-gatherer communities of the Baltic forest zone extending eastwards through Latvia into northern Russia, dating to the period 8500–5000 BC according to calibrated radiocarbon dating. It is named after the Estonian town of Kunda, about 110 kilometres (70 mi) east of Tallinn along the Gulf of Finland, near where the first extensively studied settlement was discovered on Lammasmäe Hill and in the surrounding peat bog. The oldest known settlement of the Kunda culture in Estonia is Pulli. The Kunda culture was succeeded by the Narva culture, who used pottery and showed some traces of food production.

Light skin is a human skin color that has a low level of eumelanin pigmentation as an adaptation to environments of low UV radiation. Due to migrations of people in recent centuries, light-skinned populations today are found all over the world. People with light skin pigmentation are often referred to as "white" although these usages can be ambiguous in some countries where they are used to refer specifically to certain ethnic groups or populations.

Haplogroup R1b (R-M343), previously known as Hg1 and Eu18, is a human Y-chromosome haplogroup.

Afontova Gora is a Late Upper Paleolithic and Mesolithic Siberian complex of archaeological sites located on the left bank of the Yenisey River near the city of Krasnoyarsk, Russia. Afontova Gora has cultural and genetic links to the people from Mal'ta–Buret'. The complex was first excavated in 1884 by Ivan Savenkov.

In archaeogenetics, the term Ancient North Eurasian (ANE) is the name given to an ancestral component that represents the lineage of the people of the Mal'ta–Buret' culture and populations closely related to them, such as the Upper Paleolithic individuals from Afontova Gora in Siberia. Genetic studies also revealed that the ANE are closely related to the remains of the preceding Yana Culture, which were dubbed as 'Ancient North Siberians' (ANS), and which either are directly ancestral to the ANE, or both being closely related sister lineages, sharing a common ancestral source population. The Ancient North Eurasians are deeply related to Paleolithic and Mesolithic European hunter-gatherers, but also derive a significant amount of their ancestry from a deep East Eurasian source, which they received in Siberia. Their 'Ancient West Eurasian' ancestry is represented by a lineage closer to Kostenki-14, while their 'Ancient East Eurasian' ancestry is represented by a lineage closer to the Tianyuan man.
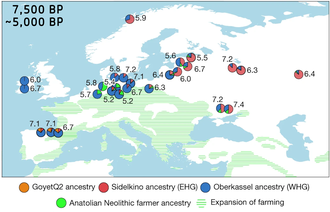
Early European Farmers (EEF), First European Farmers, Neolithic European Farmers, Ancient Aegean Farmers, or Anatolian Neolithic Farmers are names used to describe a distinct group of early Neolithic farmers who brought agriculture to Europe and Northwest Africa (Maghreb). Although the spread of agriculture from the Middle East to Europe has long been recognised through archaeology, it is only recent advances in archaeogenetics that have confirmed that this spread was strongly correlated with a migration of these farmers, and was not just a cultural exchange.

In archaeogenetics, the term Western Hunter-Gatherer is the name given to a distinct ancestral component of modern Europeans, representing descent from a population of Mesolithic hunter-gatherers who scattered over Western, Southern and Central Europe, from the British Isles in the west to the Carpathians in the east, following the retreat of the ice sheet of the Last Glacial Maximum. It is closely associated and sometimes considered synonymous with the concept of the Villabruna cluster, named after Ripari Villabruna cave in Italy, known from the terminal Pleistocene of Europe, which is largely ancestral to later WHG populations.

Caucasus hunter-gatherer (CHG), also called Satsurblia cluster, is an anatomically modern human genetic lineage, first identified in a 2015 study, based on the population genetics of several modern Western Eurasian populations.

In archaeogenetics, the term Eastern Hunter-Gatherer (EHG), sometimes East European Hunter-Gatherer, or Eastern European Hunter-Gatherer is the name given to a distinct ancestral component that represents Mesolithic hunter-gatherers of Eastern Europe.
Basal Eurasian is a proposed lineage of anatomically modern humans with reduced, or zero, archaic hominin (Neanderthal) admixture compared to other ancient non-Africans. Basal Eurasians represent a sister lineage to other Eurasians and may have originated from the Southern Middle East, specifically the Arabian peninsula, or North Africa, and are said to have contributed ancestry to various West Eurasian, South Asian, and Central Asian as well as African groups. This Basal Eurasian component is also proposed to explain the lower archaic admixture among modern West Eurasians compared to with East Eurasians, although alternatives without the need of such Basal admixture exist as well.

In archaeogenetics, the term Western Steppe Herders (WSH), or Western Steppe Pastoralists, is the name given to a distinct ancestral component first identified in individuals from the Chalcolithic steppe around the turn of the 5th millennium BC, subsequently detected in several genetically similar or directly related ancient populations including the Khvalynsk, Repin, Sredny Stog, and Yamnaya cultures, and found in substantial levels in contemporary European, Central Asian, South Asian and West Asian populations. This ancestry is often referred to as Yamnaya ancestry, Yamnaya-related ancestry, Steppe ancestry or Steppe-related ancestry.

Yuzhny Oleny, also Yuzhniy Oleniy, is an archaeological site located on Yuzhny Oleny island, in Lake Onega, Karelia.