
An anapsid is an amniote whose skull lacks one or more skull openings near the temples. Traditionally, the Anapsida are considered the most primitive subclass of amniotes, the ancestral stock from which Synapsida and Diapsida evolved, making anapsids paraphyletic. It is, however, doubtful that all anapsids lack temporal fenestra as a primitive trait, and that all the groups traditionally seen as anapsids truly lacked fenestra.

Synapsida is a diverse group of tetrapod vertebrates that includes all mammals and their extinct relatives. It is one of the two major clades of the group Amniota, the other being the more diverse group Sauropsida. Unlike other amniotes, synapsids have a single temporal fenestra, an opening low in the skull roof behind each eye socket, leaving a bony arch beneath each; this accounts for the name "synapsid". The distinctive temporal fenestra developed about 318 million years ago during the Late Carboniferous period, when synapsids and sauropsids diverged, but was subsequently merged with the orbit in early mammals.

Diapsids are a clade of sauropsids, distinguished from more primitive eureptiles by the presence of two holes, known as temporal fenestrae, in each side of their skulls. The earliest traditionally identified diapsids, the araeoscelidians, appeared about three hundred million years ago during the late Carboniferous period. All diapsids other than the most primitive ones in the clade Araeoscelidia are often placed into the clade Neodiapsida. The diapsids are extremely diverse, and include birds and all modern reptile groups, including turtles, which were historically thought to lie outside the group. All modern reptiles and birds are placed within the neodiapsid subclade Sauria. Although some diapsids have lost either one hole (lizards), or both holes, or have a heavily restructured skull, they are still classified as diapsids based on their ancestry. At least 17,084 species of diapsid animals are extant: 9,159 birds, and 7,925 snakes, lizards, tuatara, turtles, and crocodiles.
The quadratojugal is a skull bone present in many vertebrates, including some living reptiles and amphibians.

Mesosaurs were a group of small aquatic reptiles that lived during the early Permian period (Cisuralian), roughly 299 to 270 million years ago. Mesosaurs were the first known aquatic reptiles, having apparently returned to an aquatic lifestyle from more terrestrial ancestors. It is uncertain which and how many terrestrial traits these ancestors displayed; recent research cannot establish with confidence if the first amniotes were fully terrestrial, or only amphibious. Most authors consider mesosaurs to have been aquatic, although adult animals may have been amphibious, rather than completely aquatic, as indicated by their moderate skeletal adaptations to a semiaquatic lifestyle. Similarly, their affinities are uncertain; they may have been among the most basal sauropsids or among the most basal parareptiles.

Archosauromorpha is a clade of diapsid reptiles containing all reptiles more closely related to archosaurs rather than lepidosaurs. Archosauromorphs first appeared during the late Middle Permian or Late Permian, though they became much more common and diverse during the Triassic period.
A fenestra is any small opening or pore, commonly used as a term in the biological sciences. It is the Latin word for "window", and is used in various fields to describe a pore in an anatomical structure.

Euryapsida is a polyphyletic group of sauropsids that are distinguished by a single temporal fenestra, an opening behind the orbit, under which the post-orbital and squamosal bones articulate. They are different from Synapsida, which also have a single opening behind the orbit, by the placement of the fenestra. In synapsids, this opening is below the articulation of the post-orbital and squamosal bones. It is now commonly believed that euryapsids are in fact diapsids that lost the lower temporal fenestra. Euryapsids are usually considered entirely extinct, although turtles might be part of the sauropterygian clade while other authors disagree. Euryapsida may also be a synonym of Sauropterygia sensu lato.

Parareptilia ("near-reptiles") is an extinct subclass or clade of basal sauropsids/reptiles, typically considered the sister taxon to Eureptilia. Parareptiles first arose near the end of the Carboniferous period and achieved their highest diversity during the Permian period. Several ecological innovations were first accomplished by parareptiles among reptiles. These include the first reptiles to return to marine ecosystems (mesosaurs), the first bipedal reptiles, the first reptiles with advanced hearing systems, and the first large herbivorous reptiles. The only parareptiles to survive into the Triassic period were the procolophonoids, a group of small generalists, omnivores, and herbivores. The largest family of procolophonoids, the procolophonids, rediversified in the Triassic, but subsequently declined and became extinct by the end of the period.

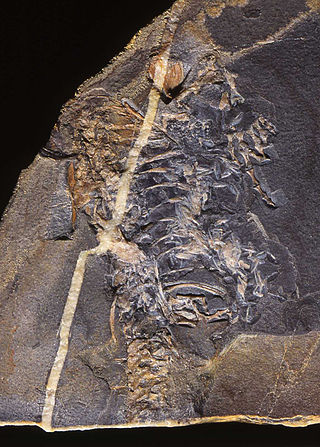
Limnoscelis was a genus of large diadectomorph tetrapods from the Late Carboniferous to early Permian of western North America. It includes two species: the type species Limnoscelis paludis from New Mexico, and Limnoscelis dynatis from Colorado, both of which are thought to have lived concurrently. No specimens of Limnoscelis are known from outside of North America. Limnoscelis was carnivorous, and likely semiaquatic, though it may have spent a significant portion of its life on land. Limnoscelis had a combination of derived amphibian and primitive reptilian features, and its placement relative to Amniota has significant implications regarding the origins of the first amniotes.

Aerosaurus is an extinct genus within Varanopidae, a family of non-mammalian synapsids. It lived between 252-299 million years ago during the Early Permian in North America. The name comes from Latin aes (aeris) “copper” and Greek sauros “lizard,” for El Cobre Canyon in northern New Mexico, where the type fossil was found and the site of former copper mines. Aerosaurus was a small to medium-bodied carnivorous synapsid characterized by its recurved teeth, triangular lateral temporal fenestra, and extended teeth row. Two species are recognized: A. greenleeorum (1937) and A. wellesi (1981).

Heleosaurus scholtzi is an extinct species of basal synapsids, known as pelycosaurs, in the family of Varanopidae during the middle Permian. At first H. scholtzi was mistakenly classified as a diapsid. Members of this family were carnivorous and had dermal armor, and somewhat resembled monitor lizards. This family was the most geologically long lived, widespread, and diverse group of early amniotes. To date only two fossils have been found in the rocks of South Africa. One of these fossils is an aggregation of five individuals.

The skull roof or the roofing bones of the skull are a set of bones covering the brain, eyes and nostrils in bony fishes and all land-living vertebrates. The bones are derived from dermal bone and are part of the dermatocranium.

Microleter is an extinct genus of basal procolophonomorph parareptiles which lived in Oklahoma during the Early Permian period. The type and only known species is Microleter mckinzieorum. Microleter is one of several parareptile taxa described from the Richards Spur fissure fills, and can be characterized from its high tooth count, lacrimal/narial contact, short postfrontal, and slit-like temporal emargination edged by the postorbital, jugal, squamosal, and quadratojugal. Contrary to Australothyris, which had a similar phylogenetic position as a basal procolophonomorph, Microleter suggests that early parareptile evolution occurred in Laurasia and that multiple lineages developed openings or emarginations in the temporal region.
Australothyris is an extinct genus of basal procolophonomorph parareptile known from the Middle Permian of Tapinocephalus Assemblage Zone, South Africa. The type and only known species is Australothyris smithi. As the most basal member of Procolophonomorpha, Australothyris helped to contextualize the origin of this major parareptile subgroup. It has been used to support the hypotheses that procolophonomorphs originated in Gondwana and ancestrally possess temporal fenestrae, due to its large and fully enclosed temporal fenestra and South African heritage. It also possessed several unique features, including a high tooth number, long postfrontal, small interpterygoid vacuity, and a specialized interaction between the stapes and quadrate.
Reptiles arose about 320 million years ago during the Carboniferous period. Reptiles, in the traditional sense of the term, are defined as animals that have scales or scutes, lay land-based hard-shelled eggs, and possess ectothermic metabolisms. So defined, the group is paraphyletic, excluding endothermic animals like birds that are descended from early traditionally-defined reptiles. A definition in accordance with phylogenetic nomenclature, which rejects paraphyletic groups, includes birds while excluding mammals and their synapsid ancestors. So defined, Reptilia is identical to Sauropsida.

Palatodonta is an extinct genus of neodiapsid reptile known from the early Middle Triassic of the Netherlands. It was initially described in 2013 as a basal placodontiform closely related to a group of marine reptiles called placodonts, characterized by their crushing teeth and shell-like body armor. Under this interpretation, Palatodonta is transitional between placodonts and less specialized reptiles. Like placodonts, it has a row of large teeth on its palate, but while these teeth are thick and blunt in placodonts, Palatodonta has palatal teeth that are thin and pointed. A 2023 study instead classified it as a sauropterygomorph and the sister taxon to Eusaurosphargis. In other words, it is close to, but not within, Sauropterygia.
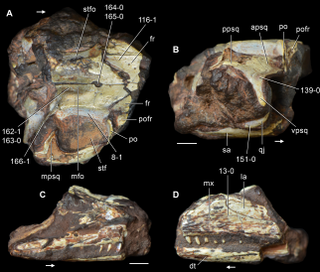
Kadimakara is an extinct genus of early archosauromorph reptile from the Arcadia Formation of Queensland, Australia. It was seemingly a very close relative of Prolacerta, a carnivorous reptile which possessed a moderately long neck. The generic name Kadimakara references prehistoric creatures from Aboriginal myths which may have been inspired by ice-age megafauna. The specific name K. australiensis relates to the fact that it was found in Australia. Prolacerta and Kadimakara were closely related to the Archosauriformes, a successful group which includes archosaurs such as crocodilians, pterosaurs, and dinosaurs.
Boreopricea is an extinct genus of archosauromorph reptile from the Early Triassic of arctic Russia. It is known from a fairly complete skeleton discovered in a borehole on Kolguyev Island, though damage to the specimen and loss of certain bones has complicated study of the genus. Boreopricea shared many similarities with various other archosauromorphs, making its classification controversial. Various studies have considered it a close relative of Prolacerta, tanystropheids, both, or neither. Boreopricea is unique among early archosauromorphs due to possessing contact between the jugal and squamosal bones at the rear half of the skull.
Eurycephalosuchus is an extinct genus of orientalosuchine alligatoroid from the Late Cretaceous Jiangxi Province of China. Known from a well preserved skull and mandible alongside various postcranial remains, Eurycephalosuchus possessed a short and broad skull with a very short skulltable. Eurycephalosuchus lived with at least one other crocodilian, an indetermined member of the clade Brevirostres. The genus is monotypic, containing only the species Eurycephalosuchus gannanensis.

