It has been suggested that this article be split out into a new article titled Neodiapsida. (Discuss) (August 2025)
Diapsids ("two arches") are a clade of sauropsids, distinguished from more primitive eureptiles by the presence of two holes, known as temporal fenestrae, in each side of their skulls. The earliest traditionally identified diapsids, the araeoscelidians, appeared about three hundred million years ago during the late Carboniferous period.[1] All diapsids other than the most primitive ones in the clade Araeoscelidia are often placed into the clade Neodiapsida. The diapsids are extremely diverse, and include birds and all modern reptile groups, including turtles, which were historically thought to lie outside the group.[2] All modern reptiles and birds are placed within the neodiapsid subclade Sauria. Although some diapsids have lost either one hole (lizards), or both holes (snakes and turtles), or have a heavily restructured skull (modern birds), they are still classified as diapsids based on their ancestry. At least 17,084 species of diapsid animals are extant: 9,159 birds,[3] and 7,925 snakes, lizards, tuatara, turtles, and crocodiles.[4]
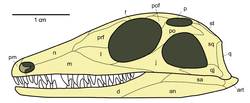
Diagram of the diapsid skull with temporal openings, unlike in anapsids
The name Diapsida means "two arches", and diapsids are traditionally classified based on their two ancestral skull openings (temporal fenestrae) posteriorly above and below the eye. This arrangement allows for the attachment of larger, stronger jaw muscles, and enables the jaw to open more widely. A more obscure ancestral characteristic is a relatively long lower arm bone (the radius) compared to the upper arm bone (humerus).
Diapsids were originally classified as one of four subclasses of the class Reptilia, all of which were based on the number and arrangement of openings in the skull. The other three subclasses were Synapsida (one opening low on the skull, for the "mammal-like reptiles"), Anapsida (no skull opening, including turtles and their relatives), and Euryapsida (one opening high on the skull, including many prehistoric marine reptiles). With the advent of phylogenetic nomenclature, this system of classification was heavily modified. Today, the synapsids are often not considered true reptiles, while Euryapsida were found to be an unnatural assemblage of diapsids that had lost one of their skull openings. Genetic studies and the discovery of the Triassic Pappochelys have shown that this is also the case in turtles, which are actually heavily modified diapsids. In phylogenetic systems, birds (descendants of traditional diapsid reptiles) are also considered to be members of this group.
Some modern studies of reptile relationships have preferred to use the name "diapsid" to refer to the crown group of all modern diapsid reptiles but not their extinct relatives. However, many researchers have also favored a more traditional definition that includes the prehistoric araeoscelidians. In 1991, Laurin defined Diapsida as a clade, "the most recent common ancestor of araeoscelidians, lepidosaurs, and archosaurs, and all its descendants".[8]
The clade Neodiapsida was given a phylogenetic definition by Laurin in 1991. He defined it as the branch-based clade containing all animals more closely related to "Younginiformes" (later, more specifically, emended to Youngina capensis) than to Petrolacosaurus (representing Araeoscelidia).[9] The earliest known neodiapsids like Orovenator are known from the Early Permian, around 290 million years ago.[10]
All genetic studies have supported the hypothesis that turtles are cladistically diapsid reptiles despite being morphologically anapsid, most placing them as more closely related to living archosaurs (including crocodiles and birds) than to lepidosaurs (lizards, snakes, etc).[11][12][13][14]
Modern reptiles and birds are placed within the neodiapsid subclade Sauria, defined as the last common ancestor of Lepidosauria (which includes lizards, snakes and the tuatara), and Archosauria (which includes crocodilians and dinosaurs, including birds, among others).[15]
A cladistic analysis by Laurin and Piñeiro (2017) recovers Parareptilia as part of Diapsida, with pareiasaurs, turtles, millerettids, and procolophonoids recovered as more derived than the basal diapsid Younginia.[16] A 2020 study by David P. Ford and Roger B. J. Benson also recovered Parareptilia as deeply nested within Diapsida as the sister group to Neodiapsida. They united this relationship between Parareptilia and Neodiapsida in the new clade Neoreptilia, defining it as the last common ancestor and all descendants of Procolophon trigoniceps and Youngina capensis.[17] However, this excludes mesosaurs, who were found to be basal among the sauropsids.[17] Other recent studies have found the more traditional arrangement of parareptiles being outside of Diapsida.[15]
↑Benton, M. J., Donoghue, P. C., Asher, R. J., Friedman, M., Near, T. J., & Vinther, J. (2015). "Constraints on the timescale of animal evolutionary history." Palaeontologia Electronica, 18.1.1FC; 1-106; palaeo-electronica.org/content/fc-1
↑Katsu, Y.; Braun, E. L.; Guillette, L. J. Jr.; Iguchi, T. (2010-03-17). "From reptilian phylogenomics to reptilian genomes: analyses of c-Jun and DJ-1 proto-oncogenes". Cytogenetic and Genome Research. 127 (2–4): 79–93. doi:10.1159/000297715. PMID20234127. S2CID12116018.
↑Bickelmann, Constanze; Müller, Johannes; and Reisz, Robert R. (2009). "The enigmatic diapsid Acerosodontosaurus piveteaui (Reptilia: Neodiapsida) from the Upper Permian of Madagascar and the paraphyly of younginiform reptiles". Canadian Journal of Earth Sciences. 49 (9): 651–661. Bibcode:2009CaJES..46..651S. doi:10.1139/E09-038.
This page is based on this Wikipedia article Text is available under the CC BY-SA 4.0 license; additional terms may apply. Images, videos and audio are available under their respective licenses.