
The Lepidosauria is a subclass or superorder of reptiles, containing the orders Squamata and Rhynchocephalia. Squamata includes lizards and snakes. Squamata contains over 9,000 species, making it by far the most species-rich and diverse order of non-avian reptiles in the present day. Rhynchocephalia was a formerly widespread and diverse group of reptiles in the Mesozoic Era. However, it is represented by only one living species: the tuatara, a superficially lizard-like reptile native to New Zealand.

Sauria is the clade containing the most recent common ancestor of Archosauria and Lepidosauria, and all its descendants. Since most molecular phylogenies recover turtles as more closely related to archosaurs than to lepidosaurs as part of Archelosauria, Sauria can be considered the crown group of diapsids, or reptiles in general. Depending on the systematics, Sauria includes all modern reptiles or most of them as well as various extinct groups.

Rhynchocephalia is an order of lizard-like reptiles that includes only one living species, the tuatara of New Zealand. Despite its current lack of diversity, during the Mesozoic rhynchocephalians were a speciose group with high morphological and ecological diversity. The oldest record of the group is dated to the Middle Triassic around 238 to 240 million years ago, and they had achieved global distribution by the Early Jurassic. Most rhynchocephalians belong to the group Sphenodontia ('wedge-teeth'). Their closest living relatives are lizards and snakes in the order Squamata, with the two orders being grouped together in the superorder Lepidosauria.

Sauropterygia is an extinct taxon of diverse, aquatic reptiles that developed from terrestrial ancestors soon after the end-Permian extinction and flourished during the Triassic before all except for the Plesiosauria became extinct at the end of that period. The plesiosaurs would continue to diversify until the end of the Mesozoic. Sauropterygians are united by a radical adaptation of their pectoral girdle, adapted to support powerful flipper strokes. Some later sauropterygians, such as the pliosaurs, developed a similar mechanism in their pelvis. It is possible that sauropterygians are a distant relatives of turtles, uniting them under the group Pantestudines, although this is still debatable as sauropterygians might be archosauromorphs or completely unrelated to both.

Neodiapsida is a clade, or major branch, of the reptilian family tree, typically defined as including all diapsids apart from some early primitive types known as the araeoscelidians. Modern reptiles and birds belong to the neodiapsid subclade Sauria.

Lepidosauromorpha is a group of reptiles comprising all diapsids closer to lizards than to archosaurs. The only living sub-group is the Lepidosauria, which contains two subdivisions, Squamata, which contains lizards and snakes, and Rhynchocephalia, the only extant species of which is the tuatara.

Younginiformes is a group of diapsid reptiles known from the Permian-Triassic of Africa and Madagascar. It has been used as a replacement for "Eosuchia". Younginiformes were historically suggested to be lepidosauromorphs, but were later suggested to be basal non-saurian neodiapsids. The group is sometimes divided into two families, Tangasauridae and Younginidae. The monophyly of the group is disputed. A 2009 study found them to be an unresolved polytomy at the base of Neodiapsida, while a 2011 study recovered the group as paraphyletic. A 2022 study recovered the Younginiformes as a monophyletic group of basal neodiapsid reptiles, also including Claudiosaurus and Saurosternon as part of the group. Some younginiforms like Hovasaurus and Acerosodontosaurus are thought to have had an amphibious lifestyle, while others like Kenyasaurus, Thadeosaurus and Youngina were probably terrestrial.

Claudiosaurus is an extinct genus of diapsid reptiles from the Late Permian Sakamena Formation of the Morondava Basin, Madagascar. It has been suggested to be semi-aquatic.

Thalattosauria is an extinct order of marine reptiles that lived in the Middle to Late Triassic. Thalattosaurs were diverse in size and shape, and are divided into two superfamilies: Askeptosauroidea and Thalattosauroidea. Askeptosauroids were endemic to the Tethys Ocean, their fossils have been found in Europe and China, and they were likely semiaquatic fish eaters with straight snouts and decent terrestrial abilities. Thalattosauroids were more specialized for aquatic life and most had unusual downturned snouts and crushing dentition. Thalattosauroids lived along the coasts of both Panthalassa and the Tethys Ocean, and were most diverse in China and western North America. The largest species of thalattosaurs grew to over 4 meters (13 feet) in length, including a long, flattened tail utilized in underwater propulsion. Although thalattosaurs bore a superficial resemblance to lizards, their exact relationships are unresolved. They are widely accepted as diapsids, but experts have variously placed them on the reptile family tree among Lepidosauromorpha, Archosauromorpha, ichthyosaurs, and/or other marine reptiles.
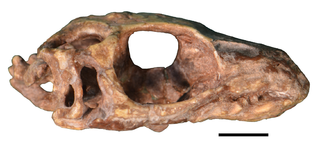
Paliguana is an extinct genus of lizard-like lepidosauromorph reptile. It is known from a single somewhat poorly preserved skull around 3 centimetres (1.2 in) in length from the earliest Triassic Katberg Formation in the upper Lystrosaurus Assemblage Zone of South Africa. It is currently the earliest known lepidosauromorph.

Acerosodontosaurus is an extinct genus of neodiapsid reptiles that lived during the Late Permian of Madagascar. The only species of Acerosodontosaurus, A. piveteaui, is known from a natural mold of a single partial skeleton including a crushed skull and part of the body and limbs. The fossil was discovered in deposits of the Lower Sakamena Formation. Based on skeletal characteristics, it has been suggested that Acerosodontosaurus individuals were at least partially aquatic.

Eunotosaurus is an extinct genus of amniote, possibly a close relative of turtles. Eunotosaurus lived in the late Middle Permian and fossils can be found in the Karoo Supergroup of South Africa. Eunotosaurus resided in the swamps of what is now southern Africa. Its ribs were wide and flat, forming broad plates similar to a primitive turtle shell, and the vertebrae were nearly identical to those of some turtles. Accordingly, it is often considered as a possible transitional fossil between turtles and their prehistoric ancestors. However, it is possible that these turtle-like features evolved independently of the same features in turtles, since other anatomical studies and phylogenetic analyses suggest that Eunotosaurus may instead have been a parareptile, an early-diverging neodiapsid unrelated to turtles, or a synapsid.

Gephyrosaurus is an extinct genus of lepidosaurian reptile known from the Late Triassic to Early Jurassic of the United Kingdom. It is generally considered to be one of the most primitive members of the clade Rhynchocephalia.
Reptiles arose about 320 million years ago during the Carboniferous period. Reptiles, in the traditional sense of the term, are defined as animals that have scales or scutes, lay land-based hard-shelled eggs, and possess ectothermic metabolisms. So defined, the group is paraphyletic, excluding endothermic animals like birds that are descended from early traditionally-defined reptiles. A definition in accordance with phylogenetic nomenclature, which rejects paraphyletic groups, includes birds while excluding mammals and their synapsid ancestors. So defined, Reptilia is identical to Sauropsida.

Marmoretta is an extinct genus of small lepidosauromorph reptile known from the Middle Jurassic (Bathonian) of Britain, as well as the Late Jurassic of Portugal. It contains a single species, Marmoretta oxoniensis.

The Erfurt Formation, also known as the Lower Keuper, is a stratigraphic formation of the Keuper group and the Germanic Trias supergroup. It was deposited during the Ladinian stage of the Triassic period. It lies above the Upper Muschelkalk and below the Middle Keuper.

Megachirella is an extinct genus of lepidosaur, possibly a stem-squamate that lived about 240 million years ago during the Middle Triassic and contains only one known species, Megachirella wachtleri. It is known from a partial skeleton discovered in the Dolomites of Northern Italy and was described in 2003.

Pantestudines or Pan-Testudines is the proposed group of all reptiles more closely related to turtles than to any other living animal. It includes both modern turtles and all of their extinct relatives. Pantestudines with a complete shell are placed in the clade Testudinata.

Vellbergia is an extinct genus of lepidosauromorph from the Middle Triassic of Germany. It contains a single species, Vellbergia bartholomaei, which is based on a tiny partial skull from the Ladinian-age Lower Keuper. Some studies have found it to be a crown group lepidosaur, closely related to Rhynchocephalia.
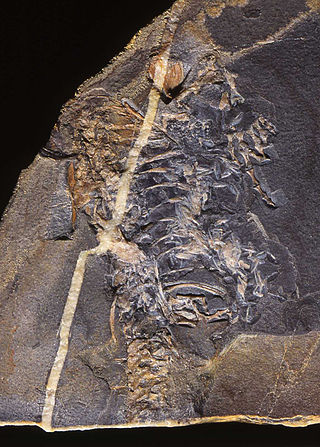
Saurosternon is an extinct genus of neodiapsid reptile from the Late Permian of South Africa. It is based on a partial skeleton split between two slabs of sandstone from the Daptocephalus Assemblage Zone. Saurosternon was one of the earliest small lizard-like reptiles to be discovered in Permian deposits of the Karoo Supergroup, preceding later discoveries such as Paliguana, Youngina, Palaeagama, and Lacertulus. The skeleton is mostly complete, though missing the head. Most of the original bone had decayed away by the time the fossil was discovered, leaving perfect molds in the sandstone slabs. What little bone remained was removed with acid by museum preparators, and the specimen was cast with latex to reconstruct the original bone shape.




