Related Research Articles
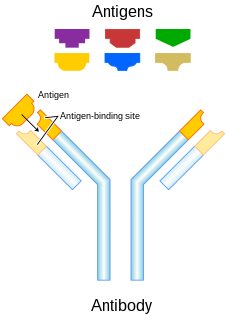
In immunology, an antigen (Ag) is a molecule or molecular structure or any foreign particulate matter or a pollen grain that can bind to a specific antibody or T-cell receptor. The presence of antigens in the body may trigger an immune response. The term antigen originally referred to a substance that is an antibody generator. Antigens can be proteins, peptides, polysaccharides, lipids, or nucleic acids.

A T cell is a type of lymphocyte. T cells are one of the important white blood cells of the immune system and play a central role in the adaptive immune response. T cells can be distinguished from other lymphocytes by the presence of a T-cell receptor (TCR) on their cell surface.

A cytotoxic T cell (also known as TC, cytotoxic T lymphocyte, CTL, T-killer cell, cytolytic T cell, CD8+ T-cell or killer T cell) is a T lymphocyte (a type of white blood cell) that kills cancer cells, cells that are infected by intracellular pathogens (such as viruses or bacteria), or cells that are damaged in other ways.

The T helper cells (Th cells), also known as CD4+ cells or CD4-positive cells, are a type of T cell that play an important role in the adaptive immune system. They aid the activity of other immune cells by releasing cytokines. They are considered essential in B cell antibody class switching, breaking cross-tolerance in dendritic cells, in the activation and growth of cytotoxic T cells, and in maximizing bactericidal activity of phagocytes such as macrophages and neutrophils. CD4+ cells are mature Th cells that express the surface protein CD4.

Natural killer cells, also known as NK cells or large granular lymphocytes (LGL), are a type of cytotoxic lymphocyte critical to the innate immune system that belong to the rapidly expanding family of known innate lymphoid cells (ILC) and represent 5–20% of all circulating lymphocytes in humans. The role of NK cells is analogous to that of cytotoxic T cells in the vertebrate adaptive immune response. NK cells provide rapid responses to virus-infected cell and other intracellular pathogens acting at around 3 days after infection, and respond to tumor formation. Typically, immune cells detect the major histocompatibility complex (MHC) presented on infected cell surfaces, triggering cytokine release, causing the death of the infected cell by lysis or apoptosis. NK cells are unique, however, as they have the ability to recognize and kill stressed cells in the absence of antibodies and MHC, allowing for a much faster immune reaction. They were named "natural killers" because of the notion that they do not require activation to kill cells that are missing "self" markers of MHC class 1. This role is especially important because harmful cells that are missing MHC I markers cannot be detected and destroyed by other immune cells, such as T lymphocyte cells.
Anergy is a term in immunobiology that describes a lack of reaction by the body's defense mechanisms to foreign substances, and consists of a direct induction of peripheral lymphocyte tolerance. An individual in a state of anergy often indicates that the immune system is unable to mount a normal immune response against a specific antigen, usually a self-antigen. Lymphocytes are said to be anergic when they fail to respond to their specific antigen. Anergy is one of three processes that induce tolerance, modifying the immune system to prevent self-destruction.

Transplant rejection occurs when transplanted tissue is rejected by the recipient's immune system, which destroys the transplanted tissue. Transplant rejection can be lessened by determining the molecular similitude between donor and recipient and by use of immunosuppressant drugs after transplant.

Superantigens (SAgs) are a class of antigens that result in excessive activation of the immune system. Specifically it causes non-specific activation of T-cells resulting in polyclonal T cell activation and massive cytokine release. SAgs are produced by some pathogenic viruses and bacteria most likely as a defense mechanism against the immune system. Compared to a normal antigen-induced T-cell response where 0.0001-0.001% of the body's T-cells are activated, these SAgs are capable of activating up to 20% of the body's T-cells. Furthermore, Anti-CD3 and Anti-CD28 antibodies (CD28-SuperMAB) have also shown to be highly potent superantigens.

A Fc receptor is a protein found on the surface of certain cells – including, among others, B lymphocytes, follicular dendritic cells, natural killer cells, macrophages, neutrophils, eosinophils, basophils, human platelets, and mast cells – that contribute to the protective functions of the immune system. Its name is derived from its binding specificity for a part of an antibody known as the Fc region. Fc receptors bind to antibodies that are attached to infected cells or invading pathogens. Their activity stimulates phagocytic or cytotoxic cells to destroy microbes, or infected cells by antibody-mediated phagocytosis or antibody-dependent cell-mediated cytotoxicity. Some viruses such as flaviviruses use Fc receptors to help them infect cells, by a mechanism known as antibody-dependent enhancement of infection.
Co-stimulation is a secondary signal which immune cells rely on to activate an immune response in the presence of an antigen-presenting cell. In the case of T cells, two stimuli are required to fully activate their immune response. During the activation of lymphocytes, co-stimulation is often crucial to the development of an effective immune response. Co-stimulation is required in addition to the antigen-specific signal from their antigen receptors.
Immune tolerance, or immunological tolerance, or immunotolerance, is a state of unresponsiveness of the immune system to substances or tissue that would otherwise have the capacity to elicit an immune response in a given organism. It is induced by prior exposure to that specific antigen and contrasts with conventional immune-mediated elimination of foreign antigens. Tolerance is classified into central tolerance or peripheral tolerance depending on where the state is originally induced—in the thymus and bone marrow (central) or in other tissues and lymph nodes (peripheral). The mechanisms by which these forms of tolerance are established are distinct, but the resulting effect is similar.
The following are notable events in the Timeline of immunology:
The following outline is provided as an overview of and topical guide to immunology:
Short Course Immune Induction Therapy or SCIIT, is a therapeutic strategy employing rapid, specific, short term-modulation of the immune system using a therapeutic agent to induce T-cell non-responsiveness, also known as operational tolerance. As an alternative strategy to immunosuppression and antigen-specific tolerance inducing therapies, the primary goal of SCIIT is to re-establish or induce peripheral immune tolerance in the context of autoimmune disease and transplant rejection through the use of biological agents. In recent years, SCIIT has received increasing attention in clinical and research settings as an alternative to immunosuppressive drugs currently used in the clinic, drugs which put the patients at risk of developing infection, cancer, and cardiovascular disease.
Alloantigen recognition refers to immune system recognition of genetically encoded polymorphisms among the genetically distinguishable members of same species. Post-transplant recognition of alloantigens occurs in secondary lymphoid organs. Donor specific antigens are recognized by recipient’s T lymphocytes and triggers adaptive pro-inflammatory response which consequently leads to rejection of allogenic transplants. Allospecific T lymphocytes may be stimulated by three major pathways: direct recognition, indirect recognition or semidirect recognition. The pathway involved in specific cases is dictated by intrinsic and extrinsic factors of allograft and directly influence nature and magnitude of T lymphocytes mediated immune response. Furthermore, variant tissues and organs such as skin or cornea or solid organ transplants can be recognized in different pathways and therefore are rejected in different fashion.

The danger model is a theory of how the immune system works. It is based on the idea that the immune system does not distinguish between self and non-self, but rather between things that might cause damage and things that will not.
Immunology is the study of the immune system during health and disease. Below is a list of immunology-related articles.
Type 1 regulatory cells or Tr1 (TR1) cells are a class of regulatory T cells participating in peripheral immunity as a subsets of CD4+ T cells. Tr1 cells regulate tolerance towards antigens of any origin. Tr1 cells are self or non-self antigen specific and their key role is to induce and maintain peripheral tolerance and suppress tissue inflammation in autoimmunity and graft vs. host disease.

An adaptive natural killer (NK) cell or memory-like NK cell is a specialized natural killer cell that has the potential to form immunological memory. They can be distinguished from cytotoxic NK (cNK) cells by their receptor expression profile and epigenome. Adaptive NK cells are so named for properties which they share with the adaptive immune system. Though adaptive NK cells do not possess antigen specificity, they exhibit dynamic expansions of defined cell subsets, increased proliferation and long-term persistence for up to 3 months in vivo, high IFN-γ production, potent cytotoxic activity upon ex vivo restimulation, and protective memory responses.
CD28 family receptors are a group of regulatory cell surface receptors expressed on immune cells. The CD28 family in turn is a subgroup of the immunoglobulin superfamily.
References
- ↑ Isoantigen Archived 2016-10-09 at the Wayback Machine at eMedicine Dictionary
- ↑ "Home". allaboutantibodies.com.
- 1 2 3 4 5 6 7 Sánchez-Fueyo A, Strom TB (2011), Immunologic basis of graft rejection and tolerance following transplantation of liver or other solid organs. Gastroenterology 140(1):51-64
- ↑ Seetharam A, Tiriveedhi V, Mohanakumar T (2010), Alloimmunity and autoimmunity in chronic rejection. Curr Opin Organ Transplant 15(4):531-536
- ↑ Fang Li, Mary E. Atz, Elaine F. Reed (2009), Human leukocyte antigen antibodies in chronic transplant vasculopathy-mechanisms and pathways. Curr Opin Immunol. 21(5): 557–562
- ↑ Walker WE, Goldstein DR (August 2007). "Neonatal B cells suppress innate toll-like receptor immune responses and modulate alloimmunity". J. Immunol. 179 (3): 1700–10. doi: 10.4049/jimmunol.179.3.1700 . PMID 17641036.
- ↑ Walsh PT, Strom TB, Turka LA (2004), Routes to transplant tolerance versus rejection: the role of cytokines. Immunity (20):121-131
- ↑ Korn T, Bettelli E, Gao W, Awasthi A, Jäger A, Strom TB, Oukka M, Kuchroo VK (2007), IL-21 initiates an alternative pathway to induce proinflammatory T(H)17 cells. Nature 448(7152):484-7
- 1 2 3 Villard J. (2011), The role of natural killer cells in human solid organ and tissue transplantation. J Innate Immun. 3(4): 395-402
- ↑ McNerney ME, Lee KM, Zhou P, Molinero L, Mashayekhi M, Guzior D, Sattar H, Kuppireddi S, Wang CR, Kumar V, Alegre ML (2006), Role of natural killer cell subsets in cardiac allograft rejection. Am J Transplant. 6(3):505-13
- ↑ Yu G, Xu X, Vu MD, Kilpatrick ED, Li XC (2006), NK cells promote transplant tolerance by killing donor antigen-presenting cells. J Exp Med. 203(8):1851-8
- ↑ De Maria A, Fogli M, Mazza S, Basso M, Picciotto A, Costa P, Congia S, Mingari MC, Moretta L (2007), Increased natural cytotoxicity receptor expression and relevant IL-10 production in NK cells from chronically infected viremic HCV patiens. Eur J Immunol. 37(2):445-55
- ↑ Lafferty KJ, Prowse SJ, Simeonovic CJ, Warren HS (1983), Immunobiology of tissue transplantation: a return to the passenger leukocyte concept. Annu Rev Immunol.1:143-73 – according to Archbold JK, Ely LK, Kjer-Nielsen L, Burrows SR, Rossjohn J, McCluskey J, Macdonald WA (2008), T-cell allorecognition and MHC-restriction – A case of Jekyll and Hyde? Mol Immunol. 45(3):583-98
- ↑ Fangmann J, Dalchau R, Fabre JW (1992), Rejection of skin allografts by indirect allorecognition of donor class I major histocompatibility complex peptides. J Exp Med. 175(6):1521-9
- ↑ Gould DS, Auchincloss H Jr (1999), Direct and indirect recognition: the role of MHC antigens in graft rejection. Immunol Today. 20(2):77-82
- ↑ Li XC, Rothstein DM, Sayegh MH (2009), Costimulatory pathways in transplantation: challenges and new developments. Immunol Rev. 229(1):271-93
- ↑ Jenkins MK, Taylor PS, Norton SD, Urdahl KB (1991), CD28 delivers a costimulatory signal involved in antigen-specific IL-2 production by human T cells. J Immunol. 147(8):2461-6 – according to Priyadharshini B, Greiner DL, Brehm MA (2012), T-cell activation and transplantation tolerance. Transplant Rev (Orlando). 26(3):212-22
- ↑ Walunas TL, Lenschow DJ, Bakker CY, Linsley PS, Freeman GJ, Green JM, Thompson CB, Bluestone JA (1994), CTLA-4 can function as a negative regulator of T cell activation. Immunity. ;1(5):405-13 – according to Priyadharshini B, Greiner DL, Brehm MA (2012), T-cell activation and transplantation tolerance. Transplant Rev (Orlando). 26(3):212-22
- ↑ Jenkins MK, Schwartz RH (1987), Antigen presentation by chemically modified splenocytes induces antigen-specific T cell unresponsiveness in vitro and in vivo. J Exp Med. 165(2):302-19
- ↑ Curtsinger JM, Mescher MF (2010), Inflammatory cytokines as a third signal for T cell activation. Curr Opin Immunol. 22(3):333-40
- ↑ Williams MA, Tyznik AJ, Bevan MJ (2006), Interleukin-2 signals during priming are required for secondary expansion of CD8+ memory T cells. Nature. 441(7095):890-3
- ↑ Welsh RM, Selin LK (2002), No one is naive: the significance of heterologous T-cell immunity. Nat Rev Immunol. 2(6):417-26
- ↑ Ashton-Chess J, Giral M, Brouard S, Soulillou JP (2007), Spontaneous operational tolerance after immunosuppressive drug withdrawal in clinical renal allotransplantation. Transplantation. 84(10):1215-9 – according to Sánchez-Fueyo A, Strom TB (2011), Immunologic basis of graft rejection and tolerance following transplantation of liver or other solid organs. Gastroenterology 140(1):51-64
- ↑ Ahmed EB, Daniels M, Alegre ML, Chong AS (2011), Bacterial infections, alloimmunity, and transplantation tolerance. Transplant Rev (Orlando). 25(1):27-35
- ↑ Ford ML, Larsen CP (2009), Translating costimulation blockade to the clinic - lessons learned from three pathways. Immunol Rev. 229(1):294-306