Structure
Protein
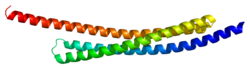
This gene encodes a 130 Å-long, arch-shaped homodimer protein. The full-length protein product spans 239 residues, 55 of which comprise the mitochondrial-targeting sequence (MTS) at its N-terminal. However, once the full-length protein is imported into the mitochondria, this sequence is excised to produce the 184-residue mature protein. [9] [10] [11] This cleavage also exposes four residues at the N-terminal, Ala-Val-Pro-Ile (AVPI), which is the core of the IAP binding domain and crucial for inhibiting XIAP. [9] [10] [11] Specifically, the tetrapeptide sequence binds the BIR3 domain of XIAP to form a stable complex between SMAC and XIAP. [9] [10] [11] The homodimer structure also facilitates SMAC-XIAP binding via the BIR2 domain, though it does not form until the protein is released into the cytoplasm as a result of outer mitochondrial membrane permeabilization. [11] Thus, monomeric SMAC mutants can still bind the BIR3 domain but not the BIR2 domain, which compromises the protein’s inhibitory function. [10] Meanwhile, mutations within the AVPI sequence lead to loss of function, though SMAC may still be able to perform IAP binding-independent functions, such as inducing the ubiquitinylation of XIAP. [10] [12]
Gene
Several alternatively spliced transcript variants that encode distinct isoforms have been described for this gene, but the validity of some transcripts, and their predicted ORFs, has not been determined conclusively. [7] [10] Two known isoforms both lack the MTS and the IAP binding domain, suggesting differential subcellular localization and function. [12]
Function
SMAC is a mitochondrial protein that promotes cytochrome c- and TNF receptor-dependent activation of apoptosis by inhibiting the effect of IAP – a group of proteins that negatively regulate apoptosis, or programmed cell death. [8] [13] SMAC is normally a mitochondrial protein localized to the mitochondrial intermembrane space, but it enters the cytosol when cells undergo apoptosis. [7] [10] [12] [14] Through the intrinsic pathway of apoptosis, BCL-2 proteins like BAK and BAX form a pore in the outer mitochondrial membrane, leading to mitochondrial membrane permeabilization and the release of both cytochrome c and SMAC. [9] [10] While cytochrome c directly activates APAF1 and caspase 9, SMAC binds IAPs, such as XIAP and cIAP proteins, to inhibit their caspase-binding activity and allow for caspase activation of apoptosis. [7] [9] [10] [12] [14] SMAC is ubiquitously expressed in many cell types, implicating it in various biological processes involving apoptosis. [15] Currently, nonapoptotic functions for SMAC remain unclear. [11]
Clinical significance
SMAC is involved in cancer, and its overexpression is linked to increased sensitivity in tumor cells to apoptosis. [7] [13] So far, SMAC overexpression has been observed to oppose cancer progression in head and neck squamous cell carcinoma, hepatocellular carcinoma, Hodgkin lymphoma, breast cancer, glioblastoma, thyroid cancer, renal cell carcinoma, testicular germ cell tumors, colorectal cancer, lung cancer, bladder cancer, endometrioid endometrial cancer, and other sarcomas. [13] [15] [16] However, the exact relationship between SMAC and leukemia and hematological diseases remains controversial. SMAC mimetics monotherapy displays improved cytotoxic effects on leukemic cell lines compared to combined therapy with other drugs, which is commonly more effective in other types of cancers. [17]
Following experimental elucidation of SMAC structure, small-molecule SMAC mimetics have been developed to mimic the tetrapeptide AVPI in the IAP binding domain of SMAC, which is responsible for binding the BIR3 domains in IAPs like XIAP, cIAP1, and cIAP2 to induce apoptosis, and sometimes, necroptosis. [9] [16] Several of the numerous SMAC mimetics designed within the last decade or so are now undergoing clinical trials, including SM-406 by Bai and colleagues and two mimetics by Genentech. These mimetics are also designed to target tumor cells directly through interacting with inflammatory proteins, such as IL-1β, which are commonly produced by solid tumor lesions. [9] Notably, preclinical studies indicate that the use of SMAC mimetics in conjunction with chemotherapy, death receptor ligands and agonists, as well as small molecule targeted drugs enhance the sensitivity of tumor cells to these treatments. [9] [13] [16] In addition to improving the success of tumor elimination, this increased sensitivity can permit smaller doses, thus minimizing side effects while maintaining efficacy. [16] Nonetheless, there still exists the potential for side effects, such as elevated levels of cytokines and chemokines in normal tissues, depending on the cellular environment. [9]
In addition to cancers, mutations in DIABLO is associated with young-adult onset of nonsyndromic deafness-64. [7]
This page is based on this
Wikipedia article Text is available under the
CC BY-SA 4.0 license; additional terms may apply.
Images, videos and audio are available under their respective licenses.