Homer protein homolog 1 or Homer1 is a neuronal protein that in humans is encoded by the HOMER1 gene. [5] [6] [7] Other names are Vesl and PSD-Zip45.
Homer protein homolog 1 or Homer1 is a neuronal protein that in humans is encoded by the HOMER1 gene. [5] [6] [7] Other names are Vesl and PSD-Zip45.
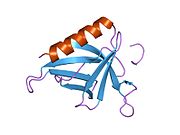
Homer1 protein has an N-terminal EVH1 domain, involved in protein interaction, and a C-terminal coiled-coil domain involved in self association. It consists of two major splice variants, short-form (Homer1a) and long-form (Homer1b and c). Homer1a has only EVH1 domain and is monomeric while Homer1b and 1c have both EVH1 and coiled-coil domains and are tetrameric. [8] [9] The coiled-coil can be further separated into N-terminal half and C-terminal half. The N-terminal half of the coiled-coil domain is predicted to be a parallel dimer while the C-terminus half is a hybrid of dimeric and anti-parallel tetrameric coiled-coil. As a whole, long Homer is predicted to have a dumbbell-like structure where two pairs of EVH1 domains are located on two sides of long (~50 nm) coiled-coil domain. [9] Mammals have Homer2 and Homer3, in addition to Homer1, which have similar domain structure. They also have similar alternatively spliced forms.

Homer1 is expressed widely in the central nervous system as well as peripheral tissue including heart, kidney, ovary, testis, and skeletal muscle. Subcellularly in neurons, Homer1 is concentrated in postsynaptic structures and constitutes a major part of the postsynaptic density.
EVH1 domain interacts with PPXXF motif. This sequence motif exists in group 1 metabotrophic glutamate receptor (mGluR1 and mGluR5), IP3 receptors (IP3R), Shank, transient receptor potential canonical (TRPC) family channels, drebrin, oligophrenin, dynamin3, CENTG1, and ryanodin receptor. [5] [7] [10] [11] [12] [13] Through its tetrameric structure, long forms of Homer (such as Homer1b and Homer1c) are proposed to cross link different proteins. For example, group 1 mGluR is crossed linked with its signaling downstream, IP3 receptor. [10] Also, through crosslinking another multimeric protein Shank, it is proposed to comprise a core of the postsynaptic density. [9]
Notably, the expression of Homer1a is induced by neuronal activity while that of Homer1b and 1c are constitutive. Thus Homer1a is classified as an immediate early gene. Homer1a, acts as a natural dominant negative form that blocks interaction between long-forms and their ligand proteins by competing with the EVH1 binding site on the ligand proteins. In this way, the short form of Homer uncouples mGluR signaling and also shrinks dendritic spine structure. [6] [14] Therefore, the short form of Homer is considered to be a part of a mechanism of homeostatic plasticity that dampens the neuronal responsiveness when input activity is too high. The long form Homer1c plays a role in synaptic plasticity and the stabilization of synaptic changes during long-term potentiation. [15]
The coiled-coil domain is reported to interact with syntaxin13 and activated Cdc42. The interaction with Cdc42 inhibit the activity of Cdc42 to remodel dendritic spine structure.
Homer1a switches mGluR5 signaling to increase AMPA receptor activity for the rapid antidepressant actions of sleep deprivation. [16]
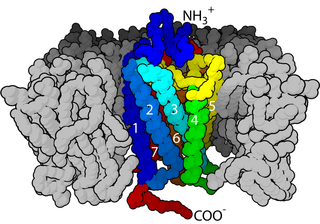
A neurotransmitter receptor is a membrane receptor protein that is activated by a neurotransmitter. Chemicals on the outside of the cell, such as a neurotransmitter, can bump into the cell's membrane, in which there are receptors. If a neurotransmitter bumps into its corresponding receptor, they will bind and can trigger other events to occur inside the cell. Therefore, a membrane receptor is part of the molecular machinery that allows cells to communicate with one another. A neurotransmitter receptor is a class of receptors that specifically binds with neurotransmitters as opposed to other molecules.

The α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor is an ionotropic transmembrane receptor for glutamate (iGluR) that mediates fast synaptic transmission in the central nervous system (CNS). It has been traditionally classified as a non-NMDA-type receptor, along with the kainate receptor. Its name is derived from its ability to be activated by the artificial glutamate analog AMPA. The receptor was first named the "quisqualate receptor" by Watkins and colleagues after a naturally occurring agonist quisqualate and was only later given the label "AMPA receptor" after the selective agonist developed by Tage Honore and colleagues at the Royal Danish School of Pharmacy in Copenhagen. The GRIA2-encoded AMPA receptor ligand binding core was the first glutamate receptor ion channel domain to be crystallized.
An inhibitory postsynaptic potential (IPSP) is a kind of synaptic potential that makes a postsynaptic neuron less likely to generate an action potential. IPSPs were first investigated in motorneurons by David P. C. Lloyd, John Eccles and Rodolfo Llinás in the 1950s and 1960s. The opposite of an inhibitory postsynaptic potential is an excitatory postsynaptic potential (EPSP), which is a synaptic potential that makes a postsynaptic neuron more likely to generate an action potential. IPSPs can take place at all chemical synapses, which use the secretion of neurotransmitters to create cell to cell signalling. Inhibitory presynaptic neurons release neurotransmitters that then bind to the postsynaptic receptors; this induces a change in the permeability of the postsynaptic neuronal membrane to particular ions. An electric current that changes the postsynaptic membrane potential to create a more negative postsynaptic potential is generated, i.e. the postsynaptic membrane potential becomes more negative than the resting membrane potential, and this is called hyperpolarisation. To generate an action potential, the postsynaptic membrane must depolarize—the membrane potential must reach a voltage threshold more positive than the resting membrane potential. Therefore, hyperpolarisation of the postsynaptic membrane makes it less likely for depolarisation to sufficiently occur to generate an action potential in the postsynaptic neurone.
In neurophysiology, long-term depression (LTD) is an activity-dependent reduction in the efficacy of neuronal synapses lasting hours or longer following a long patterned stimulus. LTD occurs in many areas of the CNS with varying mechanisms depending upon brain region and developmental progress.

Kainate receptors, or kainic acid receptors (KARs), are ionotropic receptors that respond to the neurotransmitter glutamate. They were first identified as a distinct receptor type through their selective activation by the agonist kainate, a drug first isolated from the algae Digenea simplex. They have been traditionally classified as a non-NMDA-type receptor, along with the AMPA receptor. KARs are less understood than AMPA and NMDA receptors, the other ionotropic glutamate receptors. Postsynaptic kainate receptors are involved in excitatory neurotransmission. Presynaptic kainate receptors have been implicated in inhibitory neurotransmission by modulating release of the inhibitory neurotransmitter GABA through a presynaptic mechanism.

The metabotropic glutamate receptors, or mGluRs, are a type of glutamate receptor that are active through an indirect metabotropic process. They are members of the group C family of G-protein-coupled receptors, or GPCRs. Like all glutamate receptors, mGluRs bind with glutamate, an amino acid that functions as an excitatory neurotransmitter.

Glutamate receptors are synaptic and non synaptic receptors located primarily on the membranes of neuronal and glial cells. Glutamate is abundant in the human body, but particularly in the nervous system and especially prominent in the human brain where it is the body's most prominent neurotransmitter, the brain's main excitatory neurotransmitter, and also the precursor for GABA, the brain's main inhibitory neurotransmitter. Glutamate receptors are responsible for the glutamate-mediated postsynaptic excitation of neural cells, and are important for neural communication, memory formation, learning, and regulation.
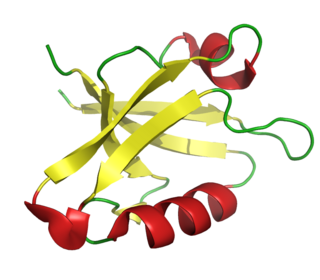
The PDZ domain is a common structural domain of 80-90 amino-acids found in the signaling proteins of bacteria, yeast, plants, viruses and animals. Proteins containing PDZ domains play a key role in anchoring receptor proteins in the membrane to cytoskeletal components. Proteins with these domains help hold together and organize signaling complexes at cellular membranes. These domains play a key role in the formation and function of signal transduction complexes. PDZ domains also play a highly significant role in the anchoring of cell surface receptors to the actin cytoskeleton via mediators like NHERF and ezrin.

The glutamate receptor, metabotropic 1, also known as GRM1, is a human gene which encodes the metabotropic glutamate receptor 1 (mGluR1) protein.

Metabotropic glutamate receptor 2 (mGluR2) is a protein that, in humans, is encoded by the GRM2 gene. mGluR2 is a G protein-coupled receptor (GPCR) that couples with the Gi alpha subunit. The receptor functions as an autoreceptor for glutamate, that upon activation, inhibits the emptying of vesicular contents at the presynaptic terminal of glutamatergic neurons.

Metabotropic glutamate receptor 3 (mGluR3) is an inhibitory Gi/G0-coupled G-protein coupled receptor (GPCR) generally localized to presynaptic sites of neurons in classical circuits. However, in higher cortical circuits in primates, mGluR3 are localized post-synaptically, where they strengthen rather than weaken synaptic connectivity. In humans, mGluR3 is encoded by the GRM3 gene. Deficits in mGluR3 signaling have been linked to impaired cognition in humans, and to increased risk of schizophrenia, consistent with their expanding role in cortical evolution.

Metabotropic glutamate receptor 5 is an excitatory Gq-coupled G protein-coupled receptor predominantly expressed on the postsynaptic sites of neurons. In humans, it is encoded by the GRM5 gene.

Glutamate receptor, metabotropic 6, also known as GRM6 or mGluR6, is a protein which in humans is encoded by the GRM6 gene.
Glutamate receptor 1 is a protein that in humans is encoded by the GRIA1 gene.

Glutamate receptor, ionotropic, kainate 1, also known as GRIK1, is a protein that in humans is encoded by the GRIK1 gene.
SH3 and multiple ankyrin repeat domains protein 2 is a protein that in humans is encoded by the SHANK2 gene. Two alternative splice variants, encoding distinct isoforms, are reported. Additional splice variants exist but their full-length nature has not been determined.

Homer protein homolog 3 is a protein that in humans is encoded by the HOMER3 gene.

Homer protein homolog 2 is a protein that in humans is encoded by the HOMER2 gene.

Activity-regulated cytoskeleton-associated protein is a plasticity protein that in humans is encoded by the ARC gene. The gene is believed to derive from a retrotransposon. The protein is found in the neurons of tetrapods and other animals where it can form virus-like capsids that transport RNA between neurons.

LY-487,379 is a drug used in scientific research that acts as a selective positive allosteric modulator for the metabotropic glutamate receptor group II subtype mGluR2. It is used to study the structure and function of this receptor subtype, and LY-487,379 along with various other mGluR2/3 agonists and positive modulators are being investigated as possible antipsychotic and anxiolytic drugs.
PDB gallery | |
|---|---|
|