Related Research Articles
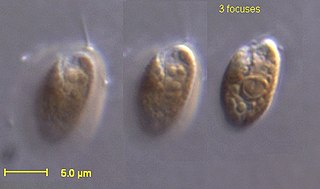
The cryptomonads are a group of algae, most of which have plastids. They are common in freshwater, and also occur in marine and brackish habitats. Each cell is around 10–50 μm in size and flattened in shape, with an anterior groove or pocket. At the edge of the pocket there are typically two slightly unequal flagella.

The centrohelids or centroheliozoa are a large group of heliozoan protists. They include both mobile and sessile forms, found in freshwater and marine environments, especially at some depth.

Chromista is a proposed but polyphyletic biological kingdom, refined from the Chromalveolata, consisting of single-celled and multicellular eukaryotic species that share similar features in their photosynthetic organelles (plastids). It includes all eukaryotes whose plastids contain chlorophyll c and are surrounded by four membranes. If the ancestor already possessed chloroplasts derived by endosymbiosis from red algae, all non-photosynthetic Chromista have secondarily lost the ability to photosynthesise. Its members might have arisen independently as separate evolutionary groups from the last eukaryotic common ancestor.

A bikont is any of the eukaryotic organisms classified in the group Bikonta. Many single-celled and multi-celled organisms are members of the group, and these, as well as the presumed ancestor, have two flagella.

The tectofilosids are a group of filose amoebae with shells. These are composed of organic materials and sometimes collected debris, in contrast to the euglyphids, which produce shells from siliceous scales. The shell usually has a single opening, but in Amphitrema and a few other genera it has two on opposite ends. The cell itself occupies most of the shell. They are most often found on marsh plants such as Sphagnum.
Chromalveolata was a eukaryote supergroup present in a major classification of 2005, then regarded as one of the six major groups within the eukaryotes. It was a refinement of the kingdom Chromista, first proposed by Thomas Cavalier-Smith in 1981. Chromalveolata was proposed to represent the organisms descended from a single secondary endosymbiosis involving a red alga and a bikont. The plastids in these organisms are those that contain chlorophyll c.
Hatena arenicola is a species of single-celled eukaryotes discovered in 2000, and first reported in 2005. It was discovered by Japanese biologists Noriko Okamoto and Isao Inouye at the University of Tsukuba, and they gave the scientific description and formal name in 2006. The species is a flagellate, and can resemble a plant at one stage of its life, in which it carries a photosynthesizing alga inside itself, or an animal, acting as predator in another stage of its life. Researchers believe that this organism is in the process of secondary endosymbiosis, in which one organism is incorporated into another, resulting in a completely new life form.

Telonemia is a phylum of microscopic eukaryotes. They are unicellular free-living flagellates with a unique combination of cell structures, including a highly complex cytoskeleton unseen in other eukaryotes. They present characteristics similar to their sister group, the SAR supergroup, such as cortical alveoli, tripartite mastigonemes and filopodia. Together, the two lineages compose the TSAR clade. They are classified in three genera and seven species, although numerous undescribed lineages are known. They are detected in all marine and freshwater environments, where they prey on bacteria and small phytoplankton through phagotrophy.

Thaumatomonadida is an order of flagellates.
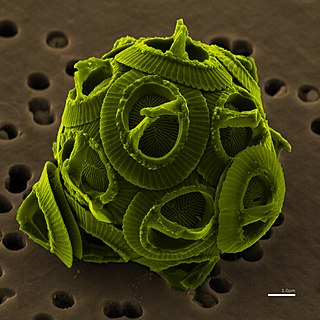
The cryptomonads-haptophytes assemblage is a proposed but disputed monophyletic grouping of unicellular eukaryotes that are not included in the SAR supergroup. Several alternative names have been used for the group, including Hacrobia ; CCTH ; and "Eukaryomonadae".
Katablepharis is a genus of single-celled eukaryotes comprising five to six species. They are heterotrophic and live in both freshwater and seawater. They have two flagella and a feeding apparatus consisting of a mouth and two arrays of microtubules.
Rhodomonas is a genus of cryptomonads. It is characterized by its red colour, the square-shaped plates of its inner periplast, its short furrow ending in a gullet, and a distinctly shaped chloroplast closely associated with its nucleomorph. Historically, Rhodomonas was characterized by its red chloroplast alone, but this no longer occurs as its taxonomy has become increasingly based on molecular and cellular data. Currently, there is some debate about the taxonomic validity of Rhodomonas as a genus and further research is needed to verify its taxonomic status. Rhodomonas is typically found in marine environments, although freshwater reports exist. It is commonly used as a live feed for various aquaculture species.
Goniomonas is a genus of Cryptomonads and contains five species. It is a genus of single-celled eukaryotes, including both freshwater and marine species. It lacks plastids, which is very unusual among all of the Cryptophyte genera. It may reflect one of only a small number of times that the Cryptophytes evolved into freshwater habitats. Goniomonas seems to have a number of freshwater relatives which have not yet been cultured and named.

Hemiselmis is a genus of cryptomonads.
Roombia truncata is a species of katablepharids, which are heterotrophic single-celled organisms.
Telonema is a genus of single-celled organisms.
Cryptista is a clade of alga-like eukaryotes. It is most likely related to Archaeplastida which includes plants and many algae, within the larger group Diaphoretickes.

Haptista is a proposed group of protists made up of centrohelids and haptophytes. Phylogenomic studies indicate that Haptista, together with Ancoracysta twista, forms a sister clade to the SAR+Telonemia supergroup, but it may also be sister to the Cryptista (+Archaeplastida). It is thus one of the earliest diverging Diaphoretickes.
Endohelea is a proposed clade of eukaryotes that are related to Archaeplastida and the SAR supergroup. They used to be considered heliozoans, but phylogenetically they belong to a group of microorganisms known as Cryptista.
Corbihelia is a proposed phylum of eukaryotes.
References
- 1 2 3 4 Okamoto N, Chantangsi C, Horák A, Leander BS, Keeling PJ (2009). Stajich JE (ed.). "Molecular Phylogeny and Description of the Novel Katablepharid Roombia truncata gen. et sp. nov., and Establishment of the Hacrobia Taxon nov". PLOS ONE. 4 (9): e7080. Bibcode:2009PLoSO...4.7080O. doi: 10.1371/journal.pone.0007080 . PMC 2741603 . PMID 19759916.
- ↑ Skuja, H. 1939. Beitrag zur Algenflora Lettlands II. Acta Horti Botanici Universitatis Latviensis 11/12: 41-168
- ↑ Clay, B. & Kugrens, P. 1999. Systematics of the Enigmatic Kathablepharids, Including EM Characterization of the Type Species, Kathablepharis phoenikoston, and New Observations on K. remigera comb. nov., Protist, Volume 150, pages 43-59, ISSN 1434-4610,
- ↑ Vørs N (1992) Ultrastructure and autecology of the marine, heterotrophic flagellate Leucocryptos marina (Braarud) Butcher 1967 (Katablepharidaceae/Kathablepharidae), with a discussion of the genera Leucocryptos and Katablepharis/Kathablepharis. Eur J Protistol 28(4): 369–389.
- ↑ Reeb, Vc; Peglar, Mt; Yoon, Hs; Bai Jr; Wu, M; Siu, P; Grafenberg, Jl; Reyes-Prieto, A; Rümmele, Se; Gross, J; Bhattacharya, D (May 2009). "Interrelationships of chromalveolates within a broadly sampled tree of photosynthetic protists". Molecular Phylogenetics & Evolution. 53 (1): 202–11. doi:10.1016/j.ympev.2009.04.012. ISSN 1055-7903. PMID 19398025.
- ↑ Noriko Okamoto; Isao Inouye (2005). "The Katablepharids are a Distant Sister Group of the Cryptophyta: A Proposal for Katablepharidophyta Divisio Nova/Kathablepharida Phylum Novum Based on SSU rDNA and Beta-Tubulin Phylogeny". Protist. 156 (2): 163–79. doi: 10.1016/j.protis.2004.12.003 . PMID 16171184.
- 1 2 Kim, E; Graham, Le (Jul 2008). Redfield, Rosemary Jeanne (ed.). "EEF2 analysis challenges the monophyly of Archaeplastida and Chromalveolata". PLOS ONE. 3 (7): e2621. Bibcode:2008PLoSO...3.2621K. doi: 10.1371/journal.pone.0002621 . PMC 2440802 . PMID 18612431.
- ↑ Cavalier-Smith; Chao; Lewis (2015), "Multiple origins of Heliozoa from flagellate ancestors: New cryptist subphylum Corbihelia, superclass Corbistoma, and monophyly of Haptista, Cryptista, Hacrobia and Chromista", Molecular Phylogenetics and Evolution, 93: 331–362, doi: 10.1016/j.ympev.2015.07.004 , PMID 26234272