
Ginkgoales are a gymnosperm order containing only one extant species: Ginkgo biloba, the ginkgo tree. The order has a long fossil record extending back to the Early Permian around 300 million years ago from fossils found worldwide. The order was a common component of Permian and Triassic flora before the super dominance of conifers.

The gymnosperms are a group of woody, perennial seed-producing plants, typically lacking the protective outer covering which surrounds the seeds in flowering plants, that include conifers, cycads, Ginkgo, and gnetophytes, forming the clade Gymnospermae The term gymnosperm comes from the composite word in Greek: γυμνόσπερμος, and literally means 'naked seeds'. The name is based on the unenclosed condition of their seeds. The non-encased condition of their seeds contrasts with the seeds and ovules of flowering plants (angiosperms), which are enclosed within an ovary. Gymnosperm seeds develop either on the surface of scales or leaves, which are often modified to form cones, or on their own as in yew, Torreya, and Ginkgo.

Paleobotany, also spelled as palaeobotany, is the branch of botany dealing with the recovery and identification of plant remains from geological contexts, and their use for the biological reconstruction of past environments (paleogeography), and the evolutionary history of plants, with a bearing upon the evolution of life in general. A synonym is paleophytology. It is a component of paleontology and paleobiology. The prefix palaeo- or paleo- means "ancient, old", and is derived from the Greek adjective παλαιός, palaios. Paleobotany includes the study of terrestrial plant fossils, as well as the study of prehistoric marine photoautotrophs, such as photosynthetic algae, seaweeds or kelp. A closely related field is palynology, which is the study of fossilized and extant spores and pollen.

Archaeopteris is an extinct genus of progymnosperm tree with fern-like leaves. A useful index fossil, this tree is found in strata dating from the Upper Devonian to Lower Carboniferous, the oldest fossils being 385 million years old, and had global distribution.

Pteridospermatophyta, also called "pteridosperms" or "seed ferns" are a polyphyletic grouping of extinct seed-producing plants. The earliest fossil evidence for plants of this type are the lyginopterids of late Devonian age. They flourished particularly during the Carboniferous and Permian periods. Pteridosperms declined during the Mesozoic Era and had mostly disappeared by the end of the Cretaceous Period, though Komlopteris seem to have survived into Eocene times, based on fossil finds in Tasmania.

Bennettitales is an extinct order of seed plants that first appeared in the Permian period and became extinct in most areas toward the end of the Cretaceous. Bennettitales were amongst the most common seed plants of the Mesozoic, and had morphologies including shrub and cycad-like forms. The foliage of bennettitaleans is superficially nearly indistinguishable from that of cycads, but they are distinguished from cycads by their more complex flower-like reproductive organs, at least some of which were likely pollinated by insects.
The Mazon Creek fossil beds are a conservation lagerstätte found near Morris, in Grundy County, Illinois. The fossils are preserved in ironstone concretions, formed approximately 309 million years ago in the mid-Pennsylvanian epoch of the Carboniferous period. These concretions frequently preserve both hard and soft tissues of animal and plant materials, as well as many soft-bodied organisms that do not normally fossilize. The quality, quantity and diversity of fossils in the area, known since the mid-nineteenth century, make the Mazon Creek lagerstätte important to paleontologists attempting to reconstruct the paleoecology of the sites. The locality was declared a National Historic Landmark in 1997.

Dioon is a genus of cycads in the family Zamiaceae. It is native to Mexico and Central America. Their habitats include tropical forests, pine-oak forest, and dry hillsides, canyons and coastal dunes.

Coal forests were the vast swathes of freshwater swamp and riparian forests that covered much of the lands on Earth's tropical regions during the late Carboniferous (Pennsylvanian) and Permian periods. As plant matter from these wetland forests decayed, enormous deposits of peat accumulated, which later became buried and converted into coal over the subsequent geologic eras.

Psaronius is an extinct genus marattialean tree fern which grew to 10m in height, and is associated with leaves of the organ genus Pecopteris and other extinct tree ferns. Originally, Psaronius was a name for the petrified stems, but today the genus is used for the entire tree fern. Psaronius tree fern fossils are found from the Carboniferous through the Permian.
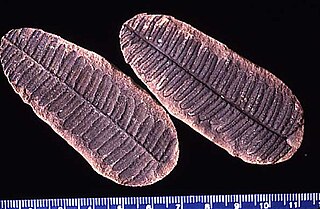
Pecopteris is a very common form genus of leaves. Most Pecopteris leaves and fronds are associated with the marattialean tree fern Psaronius. However, Pecopteris-type foliage also is borne on several filicalean ferns, and at least one seed fern. Pecopteris first appeared in the Devonian period, but flourished in the Carboniferous, especially the Pennsylvanian. Plants bearing these leaves became extinct in the Permian period, due to swamps disappearing and temperatures on Earth dropping.

The Caytoniales are an extinct order of seed plants known from fossils collected throughout the Mesozoic Era, around 252 to 66 million years ago. They are regarded as seed ferns because they are seed-bearing plants with fern-like leaves. Although at one time considered angiosperms because of their berry-like cupules, that hypothesis was later disproven. Nevertheless, some authorities consider them likely ancestors or close relatives of angiosperms. The origin of angiosperms remains unclear, and they cannot be linked with any known seed plants groups with certainty.

The Allegheny Group, often termed the Allegheny Formation, is a Pennsylvanian-age geological unit in the Appalachian Plateau. It is a major coal-bearing unit in the eastern United States, extending through western and central Pennsylvania, western Maryland and West Virginia, and southeastern Ohio. Fossils of fishes such as Bandringa are known from the Kittaning Formation, which is part of the Allegheny Group.
The Medullosales is an extinct order of pteridospermous seed plants characterised by large ovules with circular cross-section and a vascularised nucellus, complex pollen-organs, stems and rachides with a dissected stele, and frond-like leaves. Their nearest still-living relatives are the cycads.

The Lyginopteridales are an extinct group of seed plants known from the Paleozoic. They were the first plant fossils to be described as pteridosperms and, thus, the group on which the concept of pteridosperms was first developed; they are the stratigraphically oldest-known pteridosperms, occurring first in late Devonian strata; and they have the most primitive features, most notably in the structure of their ovules. They probably evolved from a group of Late Devonian progymnosperms known as the Aneurophytales, which had large, compound frond-like leaves. The Lyginopteridales became the most abundant group of pteridosperms during Mississippian times, and included both trees and smaller plants. During early and most of middle Pennsylvanian times the Medullosales took over as the more important of the larger pteridosperms but the Lyginopteridales continued to flourish as climbing (lianescent) and scrambling plants. However, later in Middle Pennsylvanian times the Lyginopteridales went into serious decline, probably being out-competed by the Callistophytales that occupied similar ecological niches but had more sophisticated reproductive strategies. A few species continued into Late Pennsylvanian times, and in Cathaysia and east equatorial Gondwana they persisted into the Late Permian, but subsequently became extinct. Most evidence of the Lyginopteridales suggests that they grew in tropical latitudes of the time, in North America, Europe and China.

Neuropteris is an extinct seed fern that existed in the Carboniferous period, known only from fossils.

Myriacantherpestes is an extinct genus of spiny millipedes from the Pennsylvanian subperiod of the Carboniferous period, known from fossils in Europe and North America.
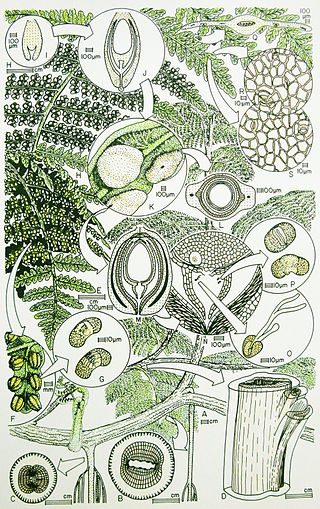
Callistophytales is an extinct order of spermatophytes which lived from the Pennsylvanian to Permian periods. They were mainly scrambling and lianescent (vine-like) plants found in the wetland "coal swamps" of Euramerica and Cathaysia. Like many other early spermatophytes, they could be described as "seed ferns", combining ovule-based reproduction with pinnate leaves superficially similar to modern ferns.
The Callistophytaceae was a family of seed ferns (pteridosperms) from the Carboniferous and Permian periods. They first appeared in late Middle Pennsylvanian (Moscovian) times, 306.5–311.7 million years ago (Ma) in the tropical coal forests of Euramerica, and became an important component of Late Pennsylvanian vegetation of clastic soils and some peat soils. The best known callistophyte was documented from Late Pennsylvanian coal ball petrifactions in North America.

Asterotheca is a genus of seedless, spore-bearing, vascularized ferns dating from the Carboniferous of the Paleozoic to the Triassic of the Mesozoic.








