
A mycorrhiza is a symbiotic association between a fungus and a plant. The term mycorrhiza refers to the role of the fungus in the plant's rhizosphere, the plant root system and its surroundings. Mycorrhizae play important roles in plant nutrition, soil biology, and soil chemistry.

An endophyte is an endosymbiont, often a bacterium or fungus, that lives within a plant for at least part of its life cycle without causing apparent disease. Endophytes are ubiquitous and have been found in all species of plants studied to date; however, most of the endophyte/plant relationships are not well understood. Some endophytes may enhance host growth and nutrient acquisition and improve the plant's ability to tolerate abiotic stresses, such as drought, and decrease biotic stresses by enhancing plant resistance to insects, pathogens and herbivores. Although endophytic bacteria and fungi are frequently studied, endophytic archaea are increasingly being considered for their role in plant growth promotion as part of the core microbiome of a plant.

An arbuscular mycorrhiza (AM) is a type of mycorrhiza in which the symbiont fungus penetrates the cortical cells of the roots of a vascular plant forming arbuscules. Arbuscular mycorrhiza is a type of endomycorrhiza along with ericoid mycorrhiza and orchid mycorrhiza. They are characterized by the formation of unique tree-like structures, the arbuscules. In addition, globular storage structures called vesicles are often encountered.
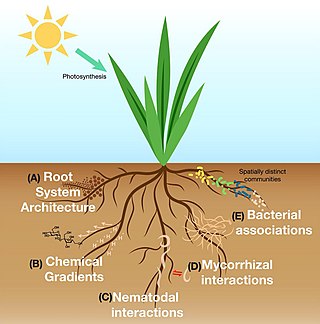
The rhizosphere is the narrow region of soil or substrate that is directly influenced by root secretions and associated soil microorganisms known as the root microbiome. Soil pores in the rhizosphere can contain many bacteria and other microorganisms that feed on sloughed-off plant cells, termed rhizodeposition, and the proteins and sugars released by roots, termed root exudates. This symbiosis leads to more complex interactions, influencing plant growth and competition for resources. Much of the nutrient cycling and disease suppression by antibiotics required by plants occurs immediately adjacent to roots due to root exudates and metabolic products of symbiotic and pathogenic communities of microorganisms. The rhizosphere also provides space to produce allelochemicals to control neighbours and relatives.

Myco-heterotrophy is a symbiotic relationship between certain kinds of plants and fungi, in which the plant gets all or part of its food from parasitism upon fungi rather than from photosynthesis. A myco-heterotroph is the parasitic plant partner in this relationship. Myco-heterotrophy is considered a kind of cheating relationship and myco-heterotrophs are sometimes informally referred to as "mycorrhizal cheaters". This relationship is sometimes referred to as mycotrophy, though this term is also used for plants that engage in mutualistic mycorrhizal relationships.
Rhizobacteria are root-associated bacteria that can have a detrimental, neutral or beneficial effect on plant growth. The name comes from the Greek rhiza, meaning root. The term usually refers to bacteria that form symbiotic relationships with many plants (mutualism). Rhizobacteria are often referred to as plant growth-promoting rhizobacteria, or PGPRs. The term PGPRs was first used by Joseph W. Kloepper in the late 1970s and has become commonly used in scientific literature.
Microbial inoculants, also known as soil inoculants or bioinoculants, are agricultural amendments that use beneficial rhizosphericic or endophytic microbes to promote plant health. Many of the microbes involved form symbiotic relationships with the target crops where both parties benefit (mutualism). While microbial inoculants are applied to improve plant nutrition, they can also be used to promote plant growth by stimulating plant hormone production. Although bacterial and fungal inoculants are common, inoculation with archaea to promote plant growth is being increasingly studied.

Fungivory or mycophagy is the process of organisms consuming fungi. Many different organisms have been recorded to gain their energy from consuming fungi, including birds, mammals, insects, plants, amoebas, gastropods, nematodes, bacteria and other fungi. Some of these, which only eat fungi, are called fungivores whereas others eat fungi as only part of their diet, being omnivores.
The mycorrhizosphere is the region around a mycorrhizal fungus in which nutrients released from the fungus increase the microbial population and its activities. The roots of most terrestrial plants, including most crop plants and almost all woody plants, are colonized by mycorrhiza-forming symbiotic fungi. In this relationship, the plant roots are infected by a fungus, but the rest of the fungal mycelium continues to grow through the soil, digesting and absorbing nutrients and water and sharing these with its plant host. The fungus in turn benefits by receiving photosynthetic sugars from its host. The mycorrhizosphere consists of roots, hyphae of the directly connected mycorrhizal fungi, associated microorganisms, and the soil in their direct influence.

A mycorrhizal network is an underground network found in forests and other plant communities, created by the hyphae of mycorrhizal fungi joining with plant roots. This network connects individual plants together. Mycorrhizal relationships are most commonly mutualistic, with both partners benefiting, but can be commensal or parasitic, and a single partnership may change between any of the three types of symbiosis at different times.

An ectomycorrhiza is a form of symbiotic relationship that occurs between a fungal symbiont, or mycobiont, and the roots of various plant species. The mycobiont is often from the phyla Basidiomycota and Ascomycota, and more rarely from the Zygomycota. Ectomycorrhizas form on the roots of around 2% of plant species, usually woody plants, including species from the birch, dipterocarp, myrtle, beech, willow, pine and rose families. Research on ectomycorrhizas is increasingly important in areas such as ecosystem management and restoration, forestry and agriculture.

Rhizophagus irregularis is an arbuscular mycorrhizal fungus used as a soil inoculant in agriculture and horticulture. Rhizophagus irregularis is also commonly used in scientific studies of the effects of arbuscular mycorrhizal fungi on plant and soil improvement. Until 2001, the species was known and widely marketed as Glomus intraradices, but molecular analysis of ribosomal DNA led to the reclassification of all arbuscular fungi from Zygomycota phylum to the Glomeromycota phylum.

The root microbiome is the dynamic community of microorganisms associated with plant roots. Because they are rich in a variety of carbon compounds, plant roots provide unique environments for a diverse assemblage of soil microorganisms, including bacteria, fungi, and archaea. The microbial communities inside the root and in the rhizosphere are distinct from each other, and from the microbial communities of bulk soil, although there is some overlap in species composition.
Orchid mycorrhizae are endomycorrhizal fungi which develop symbiotic relationships with the roots and seeds of plants of the family Orchidaceae. Nearly all orchids are myco-heterotrophic at some point in their life cycle. Orchid mycorrhizae are critically important during orchid germination, as an orchid seed has virtually no energy reserve and obtains its carbon from the fungal symbiont.
Mycorrhizal amelioration of heavy metals or pollutants is a process by which mycorrhizal fungi in a mutualistic relationship with plants can sequester toxic compounds from the environment, as a form of bioremediation.
Dr. Mohamed Hijri is a biologist who studies arbuscular mycorrhizal fungi (AMF). He is a professor of biology and research at the Institut de recherche en biologie végétale at the University of Montreal.

Funneliformis mosseae is a species of fungus in the family Glomeraceae, which is an arbuscular mycorrhizal (AM) fungi that forms symbiotic relationships with plant roots. Funneliformis mosseae has a wide distribution worldwide, and can be found in North America, South America, Europe, Africa, Asia and Australia. Funneliformis are characterized by having an easily visible septum in the area of the spore base and are often cylindrical or funnel-shaped. Funneliformis mosseae similarly resembles Glomus caledonium, however the spore wall of Funneliformis mosseae contains three layers, whereas Gl. caledonium spore walls are composed of four layers. Funneliformis is an easily cultivated species which multiplies well in trap culture, along with its high distribution, F. mosseae is not considered endangered and is often used for experimental purposes when combined with another host.
The International Collection of (Vesicular) Arbuscular Mycorrhizal Fungi (INVAM) is the largest collection of living arbuscular mycorrhizal fungi (AMF) and includes Glomeromycotan species from 6 continents. Curators of INVAM acquire, grow, identify, and elucidate the biology, taxonomy, and ecology of a diversity AMF with the mission to expand availability and knowledge of these symbiotic fungi. Culturing AMF presents difficulty as these fungi are obligate biotrophs that must complete their life cycle while in association with their plant hosts, while resting spores outside of the host are vulnerable to predation and degradation. Curators of INVAM have thus developed methods to overcome these challenges to increase the availability of AMF spores. The inception of this living collection of germplasm occurred in the 1980s and it takes the form of fungi growing in association with plant symbionts in the greenhouse, with spores preserved in cold storage within their associated rhizosphere. AMF spores acquired from INVAM have been used extensively in both basic and applied research projects in the fields of ecology, evolutionary biology, agroecology, and in restoration. INVAM is umbrellaed under the Kansas Biological Survey at The University of Kansas, an R1 Research Institution. The Kansas Biological Survey is also home to the well-known organization Monarch Watch. INVAM is currently located within the tallgrass prairie ecoregion, and many collaborators and researchers associated with INVAM study the role of AMF in the mediation of prairie biodiversity. James Bever and Peggy Schultz are the Curator and Director of Operation team, with Elizabeth Koziol and Terra Lubin as Associate Curators.
Ambispora granatensis is an arbuscular mycorrhizal fungal species in the genus Ambispora, family Ambisporaceae. It forms spores of the acaulosporois and glomoid morphs, thus the Ambispora classification. It was discovered in Granada Spain in 2010 and has unique spore characteristics, which distinguishes the species from the others in its genus.

The common symbiosis signaling pathway (CSSP) is a signaling cascade in plants that allows them to interact with symbiotic microbes. It corresponds to an ancestral pathway that plants use to interact with arbuscular mycorrhizal fungi (AMF). It is known as "common" because different evolutionary younger symbioses also use this pathway, notably the root nodule symbiosis with nitrogen-fixing rhizobia bacteria. The pathway is activated by both Nod-factor perception, as well as by Myc-factor perception that are released from AMF. The pathway is distinguished from the pathogen recognition pathways, but may have some common receptors involved in both pathogen recognition as well as CSSP. A recent work by Kevin Cope and colleagues showed that ectomycorrhizae also uses CSSP components such as Myc-factor recognition.



