
A herbivore is an animal anatomically and physiologically adapted to eating plant material, for example foliage or marine algae, for the main component of its diet. As a result of their plant diet, herbivorous animals typically have mouthparts adapted to rasping or grinding. Horses and other herbivores have wide flat teeth that are adapted to grinding grass, tree bark, and other tough plant material.
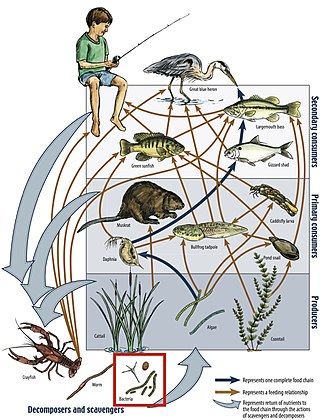
A food web is the natural interconnection of food chains and a graphical representation of what-eats-what in an ecological community. Ecologists can broadly define all life forms as either autotrophs or heterotrophs, based on their trophic levels, the position that they occupy in the food web. To maintain their bodies, grow, develop, and to reproduce, autotrophs produce organic matter from inorganic substances, including both minerals and gases such as carbon dioxide. These chemical reactions require energy, which mainly comes from the Sun and largely by photosynthesis, although a very small amount comes from bioelectrogenesis in wetlands, and mineral electron donors in hydrothermal vents and hot springs. These trophic levels are not binary, but form a gradient that includes complete autotrophs, which obtain their sole source of carbon from the atmosphere, mixotrophs, which are autotrophic organisms that partially obtain organic matter from sources other than the atmosphere, and complete heterotrophs that must feed to obtain organic matter.

Energy flow is the flow of energy through living things within an ecosystem. All living organisms can be organized into producers and consumers, and those producers and consumers can further be organized into a food chain. Each of the levels within the food chain is a trophic level. In order to more efficiently show the quantity of organisms at each trophic level, these food chains are then organized into trophic pyramids. The arrows in the food chain show that the energy flow is unidirectional, with the head of an arrow indicating the direction of energy flow; energy is lost as heat at each step along the way.

The unified neutral theory of biodiversity and biogeography is a theory and the title of a monograph by ecologist Stephen P. Hubbell. It aims to explain the diversity and relative abundance of species in ecological communities. Like other neutral theories of ecology, Hubbell assumes that the differences between members of an ecological community of trophically similar species are "neutral", or irrelevant to their success. This implies that niche differences do not influence abundance and the abundance of each species follows a random walk. The theory has sparked controversy, and some authors consider it a more complex version of other null models that fit the data better.
The metabolic theory of ecology (MTE) is the ecological component of the more general Metabolic Scaling Theory and Kleiber's law. It posits that the metabolic rate of organisms is the fundamental biological rate that governs most observed patterns in ecology. MTE is part of a larger set of theory known as metabolic scaling theory that attempts to provide a unified theory for the importance of metabolism in driving pattern and process in biology from the level of cells all the way to the biosphere.
Ecological facilitation or probiosis describes species interactions that benefit at least one of the participants and cause harm to neither. Facilitations can be categorized as mutualisms, in which both species benefit, or commensalisms, in which one species benefits and the other is unaffected. This article addresses both the mechanisms of facilitation and the increasing information available concerning the impacts of facilitation on community ecology.

Competition is an interaction between organisms or species in which both require a resource that is in limited supply. Competition lowers the fitness of both organisms involved since the presence of one of the organisms always reduces the amount of the resource available to the other.

Interspecific competition, in ecology, is a form of competition in which individuals of different species compete for the same resources in an ecosystem. This can be contrasted with mutualism, a type of symbiosis. Competition between members of the same species is called intraspecific competition.

George David Tilman, ForMemRS, is an American ecologist. He is Regents Professor and McKnight Presidential Chair in Ecology at the University of Minnesota, as well as an instructor in Conservation Biology; Ecology, Evolution, and Behavior; and Microbial Ecology. He is director of the Cedar Creek Ecosystem Science Reserve long-term ecological research station. Tilman is also a professor at University of California, Santa Barbara's Bren School of Environmental Science & Management.

In ecology, a community is a group or association of populations of two or more different species occupying the same geographical area at the same time, also known as a biocoenosis, biotic community, biological community, ecological community, or life assemblage. The term community has a variety of uses. In its simplest form it refers to groups of organisms in a specific place or time, for example, "the fish community of Lake Ontario before industrialization".

Plant ecology is a subdiscipline of ecology that studies the distribution and abundance of plants, the effects of environmental factors upon the abundance of plants, and the interactions among plants and between plants and other organisms. Examples of these are the distribution of temperate deciduous forests in North America, the effects of drought or flooding upon plant survival, and competition among desert plants for water, or effects of herds of grazing animals upon the composition of grasslands.

Universal adaptive strategy theory (UAST) is an evolutionary theory developed by J. Philip Grime in collaboration with Simon Pierce describing the general limits to ecology and evolution based on the trade-off that organisms face when the resources they gain from the environment are allocated between either growth, maintenance or regeneration – known as the universal three-way trade-off.

Intraguild predation, or IGP, is the killing and sometimes eating of a potential competitor of a different species. This interaction represents a combination of predation and competition, because both species rely on the same prey resources and also benefit from preying upon one another. Intraguild predation is common in nature and can be asymmetrical, in which one species feeds upon the other, or symmetrical, in which both species prey upon each other. Because the dominant intraguild predator gains the dual benefits of feeding and eliminating a potential competitor, IGP interactions can have considerable effects on the structure of ecological communities.
Biomass partitioning is the process by which plants divide their energy among their leaves, stems, roots, and reproductive parts. These four main components of the plant have important morphological roles: leaves take in CO2 and energy from the sun to create carbon compounds, stems grow above competitors to reach sunlight, roots absorb water and mineral nutrients from the soil while anchoring the plant, and reproductive parts facilitate the continuation of species. Plants partition biomass in response to limits or excesses in resources like sunlight, carbon dioxide, mineral nutrients, and water and growth is regulated by a constant balance between the partitioning of biomass between plant parts. An equilibrium between root and shoot growth occurs because roots need carbon compounds from photosynthesis in the shoot and shoots need nitrogen absorbed from the soil by roots. Allocation of biomass is put towards the limit to growth; a limit below ground will focus biomass to the roots and a limit above ground will favor more growth in the shoot.
The R* rule is a hypothesis in community ecology that attempts to predict which species will become dominant as the result of competition for resources. The hypothesis was formulated by American ecologist David Tilman. It predicts that if multiple species are competing for a single limiting resource, then whichever species can survive at the lowest equilibrium resource level can outcompete all other species. If two species are competing for two resources, then coexistence is only possible if each species has a lower R* on one of the resources. For example, two phytoplankton species may be able to coexist if one is more limited by nitrogen, and the other is more limited by phosphorus.
Jacob Weiner is a plant ecologist at the University of Copenhagen. Weiner has made contributions to several areas of plant ecology, including competition, allocation, allometry and application of ecological knowledge to agricultural production.
Biomass allocation is a concept in plant biology which indicates the relative proportion of plant biomass present in the different organs of a plant. It can also be used for whole plant communities.
Plant growth analysis refers to a set of concepts and equations by which changes in size of plants over time can be summarised and dissected in component variables. It is often applied in the analysis of growth of individual plants, but can also be used in a situation where crop growth is followed over time.
Herbivores' effects on plant diversity vary across environmental changes. Herbivores could increase plant diversity or decrease plant diversity. Loss of plant diversity due to climate change can also affect herbivore and plant community relationships
Plant density is the number of individual plants present per unit of ground area. It is most easily interpreted in the case of monospecific stands, where all plants belong to the same species and have germinated at the same time. However, it could also indicate the number of individual plants found at a given location.

