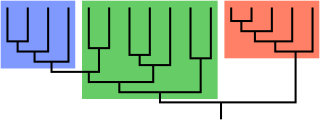
A clade, also known as a monophyletic group or natural group, is a group of organisms that are monophyletic – that is, composed of a common ancestor and all its lineal descendants – on a phylogenetic tree. Rather than the English term, the equivalent Latin term cladus is often used in taxonomical literature.

Linnaean taxonomy can mean either of two related concepts:
- The particular form of biological classification (taxonomy) set up by Carl Linnaeus, as set forth in his Systema Naturae (1735) and subsequent works. In the taxonomy of Linnaeus there are three kingdoms, divided into classes, and they, in turn, into lower ranks in a hierarchical order.
- A term for rank-based classification of organisms, in general. That is, taxonomy in the traditional sense of the word: rank-based scientific classification. This term is especially used as opposed to cladistic systematics, which groups organisms into clades. It is attributed to Linnaeus, although he neither invented the concept of ranked classification nor gave it its present form. In fact, it does not have an exact present form, as "Linnaean taxonomy" as such does not really exist: it is a collective (abstracting) term for what actually are several separate fields, which use similar approaches.

Speciation is the evolutionary process by which populations evolve to become distinct species. The biologist Orator F. Cook coined the term in 1906 for cladogenesis, the splitting of lineages, as opposed to anagenesis, phyletic evolution within lineages. Charles Darwin was the first to describe the role of natural selection in speciation in his 1859 book On the Origin of Species. He also identified sexual selection as a likely mechanism, but found it problematic.
In biology, taxonomy is the scientific study of naming, defining (circumscribing) and classifying groups of biological organisms based on shared characteristics. Organisms are grouped into taxa and these groups are given a taxonomic rank; groups of a given rank can be aggregated to form a more inclusive group of higher rank, thus creating a taxonomic hierarchy. The principal ranks in modern use are domain, kingdom, phylum, class, order, family, genus, and species. The Swedish botanist Carl Linnaeus is regarded as the founder of the current system of taxonomy, as he developed a ranked system known as Linnaean taxonomy for categorizing organisms and binomial nomenclature for naming organisms.

Apes are a clade of Old World simians native to Africa and Southeast Asia, which together with its sister group Cercopithecidae form the catarrhine clade. Apes do not have tails due to a mutation of the TBXT gene. In traditional and non-scientific use, the term "ape" can include tailless primates taxonomically considered Cercopithecidae, and is thus not equivalent to the scientific taxon Hominoidea. There are two extant branches of the superfamily Hominoidea: the gibbons, or lesser apes; and the hominids, or great apes.

A transitional fossil is any fossilized remains of a life form that exhibits traits common to both an ancestral group and its derived descendant group. This is especially important where the descendant group is sharply differentiated by gross anatomy and mode of living from the ancestral group. These fossils serve as a reminder that taxonomic divisions are human constructs that have been imposed in hindsight on a continuum of variation. Because of the incompleteness of the fossil record, there is usually no way to know exactly how close a transitional fossil is to the point of divergence. Therefore, it cannot be assumed that transitional fossils are direct ancestors of more recent groups, though they are frequently used as models for such ancestors.

A living fossil is an extant taxon that cosmetically resembles related species known only from the fossil record. To be considered a living fossil, the fossil species must be old relative to the time of origin of the extant clade. Living fossils commonly are of species-poor lineages, but they need not be. While the body plan of a living fossil remains superficially similar, it is never the same species as the remote relatives it resembles, because genetic drift would inevitably change its chromosomal structure.

This is a list of topics in evolutionary biology.
Evolutionary taxonomy, evolutionary systematics or Darwinian classification is a branch of biological classification that seeks to classify organisms using a combination of phylogenetic relationship, progenitor-descendant relationship, and degree of evolutionary change. This type of taxonomy may consider whole taxa rather than single species, so that groups of species can be inferred as giving rise to new groups. The concept found its most well-known form in the modern evolutionary synthesis of the early 1940s.

The Hominini form a taxonomic tribe of the subfamily Homininae ("hominines"). Hominini includes the extant genera Homo (humans) and Pan and in standard usage excludes the genus Gorilla (gorillas).

The tree of life or universal tree of life is a metaphor, model and research tool used to explore the evolution of life and describe the relationships between organisms, both living and extinct, as described in a famous passage in Charles Darwin's On the Origin of Species (1859).
The affinities of all the beings of the same class have sometimes been represented by a great tree. I believe this simile largely speaks the truth.
Phylogenetic nomenclature is a method of nomenclature for taxa in biology that uses phylogenetic definitions for taxon names as explained below. This contrasts with the traditional approach, in which taxon names are defined by a type, which can be a specimen or a taxon of lower rank, and a description in words. Phylogenetic nomenclature is currently regulated by the International Code of Phylogenetic Nomenclature (PhyloCode).

Dawkins vs. Gould: Survival of the Fittest is a book about the differing views of biologists Richard Dawkins and Stephen Jay Gould by philosopher of biology Kim Sterelny. When first published in 2001 it became an international best-seller. A new edition was published in 2007 to include Gould's The Structure of Evolutionary Theory finished shortly before his death in 2002, and more recent works by Dawkins. The synopsis below is from the 2007 publication.

The Hominidae, whose members are known as the great apes or hominids, are a taxonomic family of primates that includes eight extant species in four genera: Pongo ; Gorilla ; Pan ; and Homo, of which only modern humans remain.

Evolution is the process of change in all forms of life over generations, and evolutionary biology is the study of how evolution occurs. Biological populations evolve through genetic changes that correspond to changes in the organisms' observable traits. Genetic changes include mutations, which are caused by damage or replication errors in organisms' DNA. As the genetic variation of a population drifts randomly over generations, natural selection gradually leads traits to become more or less common based on the relative reproductive success of organisms with those traits.
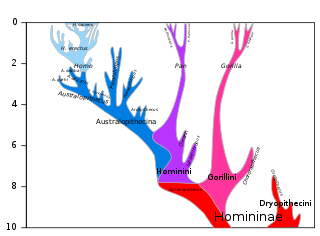
The chimpanzee–human last common ancestor (CHLCA) is the last common ancestor shared by the extant Homo (human) and Pan genera of Hominini. Due to complex hybrid speciation, it is not currently possible to give a precise estimate on the age of this ancestral population. While "original divergence" between populations may have occurred as early as 13 million years ago (Miocene), hybridization may have been ongoing until as recently as 4 million years ago (Pliocene).

The Hippocampinae are a subfamily of small marine fishes in the family Syngnathidae. Depending on the classification system used, it comprises either seahorses and pygmy pipehorses, or only seahorses.

The following outline is provided as an overview of and topical guide to evolution:
This glossary of evolutionary biology is a list of definitions of terms and concepts used in the study of evolutionary biology, population biology, speciation, and phylogenetics, as well as sub-disciplines and related fields. For additional terms from related glossaries, see Glossary of genetics, Glossary of ecology, and Glossary of biology.
The coral of life is a metaphor or a mathematical model useful to illustrate evolution of life or phylogeny at various levels of resolution, including individual organisms, populations, species and large taxonomic groups. Its use in biology resolves several practical and conceptual difficulties that are associated with the tree of life.




