Human evolution is the evolutionary process within the history of primates that led to the emergence of Homo sapiens as a distinct species of the hominid family that includes all the great apes. This process involved the gradual development of traits such as human bipedalism, dexterity, and complex language, as well as interbreeding with other hominins, indicating that human evolution was not linear but weblike. The study of the origins of humans, variously known by the terms anthropogeny, anthropogenesis, or anthropogony, involves several scientific disciplines, including physical and evolutionary anthropology, paleontology, and genetics.

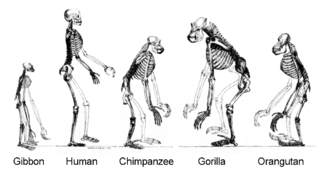
Homininae, also called "African hominids" or "African apes", is a subfamily of Hominidae. It includes two tribes, with their extant as well as extinct species: 1) the tribe Hominini ―and 2) the tribe Gorillini (gorillas). Alternatively, the genus Pan is sometimes considered to belong to its own third tribe, Panini. Homininae comprises all hominids that arose after orangutans split from the line of great apes. The Homininae cladogram has three main branches, which lead to gorillas and to humans and chimpanzees. There are two living species of Panina and two living species of gorillas, but only one extant human species. Traces of extinct Homo species, including Homo floresiensis have been found with dates as recent as 40,000 years ago. Organisms in this subfamily are described as hominine or hominines.

Homo habilis is an extinct species of archaic human from the Early Pleistocene of East and South Africa about 2.8 million years ago to 1.65 million years ago (mya). Upon species description in 1964, H. habilis was highly contested, with many researchers recommending it be synonymised with Australopithecus africanus, the only other early hominin known at the time, but H. habilis received more recognition as time went on and more relevant discoveries were made. By the 1980s, H. habilis was proposed to have been a human ancestor, directly evolving into Homo erectus which directly led to modern humans. This viewpoint is now debated. Several specimens with insecure species identification were assigned to H. habilis, leading to arguments for splitting, namely into "H. rudolfensis" and "H. gautengensis" of which only the former has received wide support.
Early modern human (EMH), or anatomically modern human (AMH), are terms used to distinguish Homo sapiens that are anatomically consistent with the range of phenotypes seen in contemporary humans, from extinct archaic human species. This distinction is useful especially for times and regions where anatomically modern and archaic humans co-existed, for example, in Paleolithic Europe. Among the oldest known remains of Homo sapiens are those found at the Omo-Kibish I archaeological site in south-western Ethiopia, dating to about 233,000 to 196,000 years ago, the Florisbad site in South Africa, dating to about 259,000 years ago, and the Jebel Irhoud site in Morocco, dated about 315,000 years ago.

Homo ergaster is an extinct species or subspecies of archaic humans who lived in Africa in the Early Pleistocene. Whether H. ergaster constitutes a species of its own or should be subsumed into H. erectus is an ongoing and unresolved dispute within palaeoanthropology. Proponents of synonymisation typically designate H. ergaster as "African Homo erectus" or "Homo erectus ergaster". The name Homo ergaster roughly translates to "working man", a reference to the more advanced tools used by the species in comparison to those of their ancestors. The fossil range of H. ergaster mainly covers the period of 1.7 to 1.4 million years ago, though a broader time range is possible. Though fossils are known from across East and Southern Africa, most H. ergaster fossils have been found along the shores of Lake Turkana in Kenya. There are later African fossils, some younger than 1 million years ago, that indicate long-term anatomical continuity, though it is unclear if they can be formally regarded as H. ergaster specimens. As a chronospecies, H. ergaster may have persisted to as late as 600,000 years ago, when new lineages of Homo arose in Africa.

Homo heidelbergensis is an extinct species or subspecies of archaic human which existed during the Middle Pleistocene. It was subsumed as a subspecies of H. erectus in 1950 as H. e. heidelbergensis, but towards the end of the century, it was more widely classified as its own species. It is debated whether or not to constrain H. heidelbergensis to only Europe or to also include African and Asian specimens, and this is further confounded by the type specimen being a jawbone, because jawbones feature few diagnostic traits and are generally missing among Middle Pleistocene specimens. Thus, it is debated if some of these specimens could be split off into their own species or a subspecies of H. erectus. Because the classification is so disputed, the Middle Pleistocene is often called the "muddle in the middle".

The Lower Paleolithic is the earliest subdivision of the Paleolithic or Old Stone Age. It spans the time from around 3.3 million years ago when the first evidence for stone tool production and use by hominins appears in the current archaeological record, until around 300,000 years ago, spanning the Oldowan and Acheulean lithics industries.

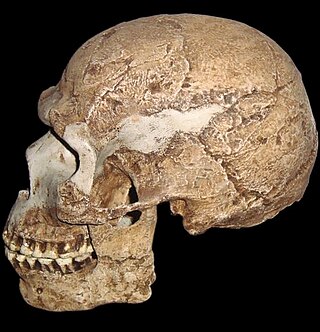
The Steinheim skull is a fossilized skull of a Homo neanderthalensis or Homo heidelbergensis found on 24 July 1933 near Steinheim an der Murr, Germany.

The Hominini form a taxonomic tribe of the subfamily Homininae ("hominines"). Hominini includes the extant genera Homo (humans) and Pan and in standard usage excludes the genus Gorilla (gorillas).
Human taxonomy is the classification of the human species within zoological taxonomy. The systematic genus, Homo, is designed to include both anatomically modern humans and extinct varieties of archaic humans. Current humans have been designated as subspecies Homo sapiens sapiens, differentiated, according to some, from the direct ancestor, Homo sapiens idaltu.
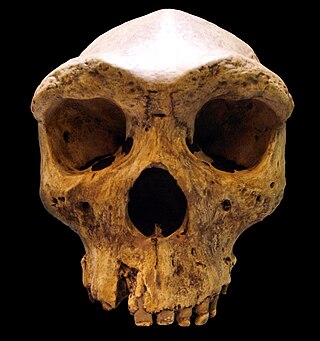
Archaic humans is a broad category denoting all species of the genus Homo that are not Homo sapiens. Among the earliest modern human remains are those from Jebel Irhoud in Morocco, Florisbad in South Africa (259 ka), and Omo-Kibish I in southern Ethiopia. Some examples of archaic humans include H. antecessor (1200–770 ka), H. bodoensis (1200–300 ka), H. heidelbergensis (600–200 ka), Neanderthals, H. rhodesiensis (300–125 ka) and Denisovans,

Homo erectus is an extinct species of archaic human from the Pleistocene, with its earliest occurrence about 2 million years ago. Its specimens are among the first recognizable members of the genus Homo.

The Hominidae, whose members are known as the great apes or hominids, are a taxonomic family of primates that includes eight extant species in four genera: Pongo ; Gorilla ; Pan ; and Homo, of which only modern humans remain.
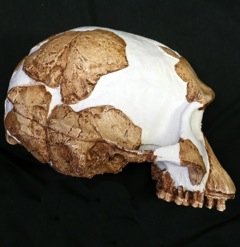
Homo gautengensis is a species name proposed by anthropologist Darren Curnoe in 2010 for South African hominin fossils otherwise attributed to H. habilis, H. ergaster, or, in some cases, Australopithecus or Paranthropus. The fossils assigned to the species by Curnoe cover a vast temporal range, from about 1.8 million years ago to potentially as late as 0.8 million years ago, meaning that if the species is considered valid, H. gautengensis would be both one of the earliest and one of the longest lived species of Homo.

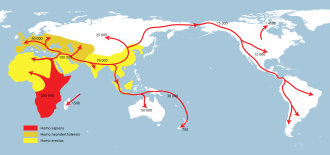
Several expansions of populations of archaic humans out of Africa and throughout Eurasia took place in the course of the Lower Paleolithic, and into the beginning Middle Paleolithic, between about 2.1 million and 0.2 million years ago (Ma). These expansions are collectively known as Out of Africa I, in contrast to the expansion of Homo sapiens (anatomically modern humans) into Eurasia, which may have begun shortly after 0.2 million years ago.
Changes to the dental morphology and jaw are major elements of hominid evolution. These changes were driven by the types and processing of food eaten. The evolution of the jaw is thought to have facilitated encephalization, speech, and the formation of the uniquely human chin.

Homo naledi is an extinct species of archaic human discovered in 2013 in the Rising Star Cave system, Gauteng province, South Africa, dating to the Middle Pleistocene 335,000–236,000 years ago. The initial discovery comprises 1,550 specimens of bone, representing 737 different skeletal elements, and at least 15 different individuals. Despite this exceptionally high number of specimens, their classification with other Homo species remains unclear.
The diet of known human ancestors varies dramatically over time. Strictly speaking, according to evolutionary anthropologists and archaeologists, there is not a single hominin Paleolithic diet. The Paleolithic covers roughly 2.8 million years, concurrent with the Pleistocene, and includes multiple human ancestors with their own evolutionary and technological adaptations living in a wide variety of environments. This fact with the difficulty of finding conclusive evidence often makes broad generalizations of the earlier human diets very difficult. Our pre-hominin primate ancestors were broadly herbivorous, relying on either foliage or fruits and nuts and the shift in dietary breadth during the Paleolithic is often considered a critical point in hominin evolution. A generalization between Paleolithic diets of the various human ancestors that many anthropologists do make is that they are all to one degree or another omnivorous and are inextricably linked with tool use and new technologies. Nonetheless, according to the California Academy of Sciences, "Prior to about 3.5 million years ago, early humans dined almost exclusively on leaves and fruits from trees, shrubs, and herbs—similar to modern-day gorillas and chimpanzees."
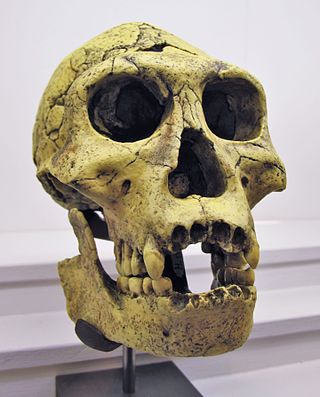
The Dmanisi hominins, Dmanisi people, or Dmanisi man were a population of Early Pleistocene hominins whose fossils have been recovered at Dmanisi, Georgia. The fossils and stone tools recovered at Dmanisi range in age from 1.85 to 1.77 million years old, making the Dmanisi hominins the earliest well-dated hominin fossils in Eurasia and the best preserved fossils of early Homo from a single site so early in time, though earlier fossils and artifacts have been found in Asia. Though their precise classification is controversial and disputed, the Dmanisi fossils are highly significant within research on early hominin migrations out of Africa. The Dmanisi hominins are known from over a hundred postcranial fossils and five famous well-preserved skulls, referred to as Dmanisi Skulls 1–5.

The archaeological site of Atapuerca is located in the province of Burgos in the north of Spain and is notable for its evidence of early human occupation. Bone fragments from around 800,000 years ago, found in its Gran Dolina cavern, provide the oldest known evidence of hominid settlement in Western Europe and of hominid cannibalism anywhere in the world.