Natural killer cells, also known as NK cells or large granular lymphocytes (LGL), are a type of cytotoxic lymphocyte critical to the innate immune system. They belong to the rapidly expanding family of known innate lymphoid cells (ILC) and represent 5–20% of all circulating lymphocytes in humans. The role of NK cells is analogous to that of cytotoxic T cells in the vertebrate adaptive immune response. NK cells provide rapid responses to virus-infected cells, stressed cells, tumor cells, and other intracellular pathogens based on signals from several activating and inhibitory receptors. Most immune cells detect the antigen presented on major histocompatibility complex I (MHC-I) on infected cell surfaces, but NK cells can recognize and kill stressed cells in the absence of antibodies and MHC, allowing for a much faster immune reaction. They were named "natural killers" because of the notion that they do not require activation to kill cells that are missing "self" markers of MHC class I. This role is especially important because harmful cells that are missing MHC I markers cannot be detected and destroyed by other immune cells, such as T lymphocyte cells.

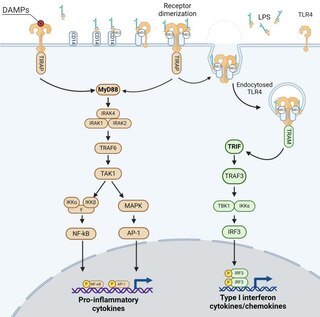
Toll-like receptors (TLRs) are a class of proteins that play a key role in the innate immune system. They are single-spanning receptors usually expressed on sentinel cells such as macrophages and dendritic cells, that recognize structurally conserved molecules derived from microbes. Once these microbes have reached physical barriers such as the skin or intestinal tract mucosa, they are recognized by TLRs, which activate immune cell responses. The TLRs include TLR1, TLR2, TLR3, TLR4, TLR5, TLR6, TLR7, TLR8, TLR9, TLR10, TLR11, TLR12, and TLR13. Humans lack genes for TLR11, TLR12 and TLR13 and mice lack a functional gene for TLR10. The receptors TLR1, TLR2, TLR4, TLR5, TLR6, and TLR10 are located on the cell membrane, whereas TLR3, TLR7, TLR8, and TLR9 are located in intracellular vesicles.
Pathogen-associated molecular patterns (PAMPs) are small molecular motifs conserved within a class of microbes, but not present in the host. They are recognized by toll-like receptors (TLRs) and other pattern recognition receptors (PRRs) in both plants and animals. This allows the innate immune system to recognize pathogens and thus, protect the host from infection.
Pattern recognition receptors (PRRs) play a crucial role in the proper function of the innate immune system. PRRs are germline-encoded host sensors, which detect molecules typical for the pathogens. They are proteins expressed mainly by cells of the innate immune system, such as dendritic cells, macrophages, monocytes, neutrophils, as well as by epithelial cells, to identify two classes of molecules: pathogen-associated molecular patterns (PAMPs), which are associated with microbial pathogens, and damage-associated molecular patterns (DAMPs), which are associated with components of host's cells that are released during cell damage or death. They are also called primitive pattern recognition receptors because they evolved before other parts of the immune system, particularly before adaptive immunity. PRRs also mediate the initiation of antigen-specific adaptive immune response and release of inflammatory cytokines.

IRAK-4, in the IRAK family, is a protein kinase involved in signaling innate immune responses from Toll-like receptors. It also supports signaling from T-cell receptors. IRAK4 contains domain structures which are similar to those of IRAK1, IRAK2, IRAKM and Pelle. IRAK4 is unique compared to IRAK1, IRAK2 and IRAKM in that it functions upstream of the other IRAKs, but is more similar to Pelle in this trait. IRAK4 has important clinical applications.

Myeloid differentiation primary response 88 (MYD88) is a protein that, in humans, is encoded by the MYD88 gene. originally discovered in the laboratory of Dan A. Liebermann as a Myeloid differentiation primary response gene.

TIR domain containing adaptor molecule 1 is an adapter in responding to activation of toll-like receptors (TLRs). It mediates the rather delayed cascade of two TLR-associated signaling cascades, where the other one is dependent upon a MyD88 adapter.

Toll-like receptor 7, also known as TLR7, is a protein that in humans is encoded by the TLR7 gene. Orthologs are found in mammals and birds. It is a member of the toll-like receptor (TLR) family and detects single stranded RNA.

Toll-like receptor 5, also known as TLR5, is a protein which in humans is encoded by the TLR5 gene. It is a member of the toll-like receptor (TLR) family. TLR5 is known to recognize bacterial flagellin from invading mobile bacteria. It has been shown to be involved in the onset of many diseases, which includes Inflammatory bowel disease. Recent studies have also shown that malfunctioning of TLR5 is likely related to rheumatoid arthritis, osteoclastogenesis, and bone loss. Abnormal TLR5 functioning is related to the onset of gastric, cervical, endometrial and ovarian cancers.

Lymphocyte antigen 96, also known as "Myeloid Differentiation factor 2 (MD-2)," is a protein that in humans is encoded by the LY96 gene.

Toll-like receptor 6 is a protein that in humans is encoded by the TLR6 gene. TLR6 is a transmembrane protein, member of toll-like receptor family, which belongs to the pattern recognition receptor (PRR) family. TLR6 acts in a heterodimer form with toll-like receptor 2 (TLR2). Its ligands include multiple diacyl lipopeptides derived from gram-positive bacteria and mycoplasma and several fungal cell wall saccharides. After dimerizing with TLR2, the NF-κB intracellular signalling pathway is activated, leading to a pro-inflammatory cytokine production and activation of innate immune response. TLR6 has also been designated as CD286.

Toll-like receptor 9 is a protein that in humans is encoded by the TLR9 gene. TLR9 has also been designated as CD289. It is a member of the toll-like receptor (TLR) family. TLR9 is an important receptor expressed in immune system cells including dendritic cells, macrophages, natural killer cells, and other antigen presenting cells. TLR9 is expressed on endosomes internalized from the plasma membrane, binds DNA, and triggers signaling cascades that lead to a pro-inflammatory cytokine response. Cancer, infection, and tissue damage can all modulate TLR9 expression and activation. TLR9 is also an important factor in autoimmune diseases, and there is active research into synthetic TLR9 agonists and antagonists that help regulate autoimmune inflammation.
Interleukin-1 receptor-associated kinase 1 (IRAK-1) is an enzyme in humans encoded by the IRAK1 gene. IRAK-1 plays an important role in the regulation of the expression of inflammatory genes by immune cells, such as monocytes and macrophages, which in turn help the immune system in eliminating bacteria, viruses, and other pathogens. IRAK-1 is part of the IRAK family consisting of IRAK-1, IRAK-2, IRAK-3, and IRAK-4, and is activated by inflammatory molecules released by signaling pathways during pathogenic attack. IRAK-1 is classified as a kinase enzyme, which regulates pathways in both innate and adaptive immune systems.

Bruce Alan Beutler is an American immunologist and geneticist. Together with Jules A. Hoffmann, he received one-half of the 2011 Nobel Prize in Physiology or Medicine, for "discoveries concerning the activation of innate immunity." Beutler discovered the long-elusive receptor for lipopolysaccharide. He did so by identifying spontaneous mutations in the gene coding for mouse Toll-like receptor 4 (Tlr4) in two unrelated strains of LPS-refractory mice and proving they were responsible for that phenotype. Subsequently, and chiefly through the work of Shizuo Akira, other TLRs were shown to detect signature molecules of most infectious microbes, in each case triggering an innate immune response.

Single Ig IL-1-related receptor (SIGIRR), also called Toll/Interleukin-1 receptor 8 (TIR8) or Interleukin-1 receptor 8 (IL-1R8), is transmembrane protein encoded by gene SIGIRR, which modulate inflammation, immune response, and tumorigenesis of colonic epithelial cells.

TIR domain-containing adapter molecule 2 is a protein that in humans is encoded by the TICAM2 gene.

The toll-interleukin-1 receptor (TIR) homology domain is an intracellular signaling domain found in MyD88, SARM1, interleukin-1 receptors, toll receptors and many plant R proteins. It contains three highly conserved regions, and mediates protein-protein interactions between the toll-like receptors (TLRs) and signal-transduction components. TIR-like motifs are also found in plant proteins where they are involved in resistance to disease and in bacteria where they are associated with virulence. When activated, TIR domains recruit cytoplasmic adaptor proteins MyD88 (UniProt Q99836) and TOLLIP (toll-interacting protein, UniProt Q9H0E2). In turn, these associate with various kinases to set off signaling cascades. Some TIR domains have also been found to have intrinsic NAD+ cleavage activity, such as in SARM1. In the case of SARM1, the TIR NADase activity leads to the production of Nam, ADPR and cADPR and the activation of downstream pathways involved in Wallerian degeneration and neuron death.
Murine caspase-11, and its human homologs caspase-4 and caspase-5, are mammalian intracellular receptor proteases activated by TLR4 and TLR3 signaling during the innate immune response. Caspase-11, also termed the non-canonical inflammasome, is activated by TLR3/TLR4-TRIF signaling and directly binds cytosolic lipopolysaccharide (LPS), a major structural element of Gram-negative bacterial cell walls. Activation of caspase-11 by LPS is known to cause the activation of other caspase proteins, leading to septic shock, pyroptosis, and often organismal death.
The interleukin-1 receptor (IL-1R) associated kinase (IRAK) family plays a crucial role in the protective response to pathogens introduced into the human body by inducing acute inflammation followed by additional adaptive immune responses. IRAKs are essential components of the Interleukin-1 receptor signaling pathway and some Toll-like receptor signaling pathways. Toll-like receptors (TLRs) detect microorganisms by recognizing specific pathogen-associated molecular patterns (PAMPs) and IL-1R family members respond the interleukin-1 (IL-1) family cytokines. These receptors initiate an intracellular signaling cascade through adaptor proteins, primarily, MyD88. This is followed by the activation of IRAKs. TLRs and IL-1R members have a highly conserved amino acid sequence in their cytoplasmic domain called the Toll/Interleukin-1 (TIR) domain. The elicitation of different TLRs/IL-1Rs results in similar signaling cascades due to their homologous TIR motif leading to the activation of mitogen-activated protein kinases (MAPKs) and the IκB kinase (IKK) complex, which initiates a nuclear factor-κB (NF-κB) and AP-1-dependent transcriptional response of pro-inflammatory genes. Understanding the key players and their roles in the TLR/IL-1R pathway is important because the presence of mutations causing the abnormal regulation of Toll/IL-1R signaling leading to a variety of acute inflammatory and autoimmune diseases.
Jonathan C. Kagan is an American immunologist and the Marian R. Neutra, Ph.D. Professor of Pediatrics at Harvard Medical School. He is also the director of Basic Research and Shwachman Chair in Gastroenterology at Boston Children's Hospital. Kagan is a world leader in defining the molecular basis of innate immunity and inflammation.