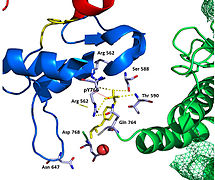
A tyrosine kinase is an enzyme that can transfer a phosphate group from ATP to the tyrosine residues of specific proteins inside a cell. It functions as an "on" or "off" switch in many cellular functions.

In cellular biology, paracrine signaling is a form of cell signaling, a type of cellular communication in which a cell produces a signal to induce changes in nearby cells, altering the behaviour of those cells. Signaling molecules known as paracrine factors diffuse over a relatively short distance, as opposed to cell signaling by endocrine factors, hormones which travel considerably longer distances via the circulatory system; juxtacrine interactions; and autocrine signaling. Cells that produce paracrine factors secrete them into the immediate extracellular environment. Factors then travel to nearby cells in which the gradient of factor received determines the outcome. However, the exact distance that paracrine factors can travel is not certain.
Fibroblast growth factors (FGF) are a family of cell signalling proteins produced by macrophages; they are involved in a wide variety of processes, most notably as crucial elements for normal development in animal cells. Any irregularities in their function lead to a range of developmental defects. These growth factors typically act as systemic or locally circulating molecules of extracellular origin that activate cell surface receptors. A defining property of FGFs is that they bind to heparin and to heparan sulfate. Thus, some are sequestered in the extracellular matrix of tissues that contains heparan sulfate proteoglycans and are released locally upon injury or tissue remodeling.

Receptor tyrosine kinases (RTKs) are the high-affinity cell surface receptors for many polypeptide growth factors, cytokines, and hormones. Of the 90 unique tyrosine kinase genes identified in the human genome, 58 encode receptor tyrosine kinase proteins. Receptor tyrosine kinases have been shown not only to be key regulators of normal cellular processes but also to have a critical role in the development and progression of many types of cancer. Mutations in receptor tyrosine kinases lead to activation of a series of signalling cascades which have numerous effects on protein expression. Receptor tyrosine kinases are part of the larger family of protein tyrosine kinases, encompassing the receptor tyrosine kinase proteins which contain a transmembrane domain, as well as the non-receptor tyrosine kinases which do not possess transmembrane domains.

Platelet-derived growth factor receptors (PDGF-R) are cell surface tyrosine kinase receptors for members of the platelet-derived growth factor (PDGF) family. PDGF subunits -A and -B are important factors regulating cell proliferation, cellular differentiation, cell growth, development and many diseases including cancer. There are two forms of the PDGF-R, alpha and beta each encoded by a different gene. Depending on which growth factor is bound, PDGF-R homo- or heterodimerizes.
The fibroblast growth factor receptors (FGFR) are, as their name implies, receptors that bind to members of the fibroblast growth factor (FGF) family of proteins. Some of these receptors are involved in pathological conditions. For example, a point mutation in FGFR3 can lead to achondroplasia.

Fibroblast growth factor receptor 2 (FGFR2) also known as CD332 is a protein that in humans is encoded by the FGFR2 gene residing on chromosome 10. FGFR2 is a receptor for fibroblast growth factor.

Fibroblast growth factor receptor 3 is a protein that in humans is encoded by the FGFR3 gene. FGFR3 has also been designated as CD333. The gene, which is located on chromosome 4, location p16.3, is expressed in tissues such as the cartilage, brain, intestine, and kidneys.
Platelet-derived growth factor receptor beta is a protein that in humans is encoded by the PDGFRB gene. Mutations in PDGFRB are mainly associated with the clonal eosinophilia class of malignancies.

Phospholipase C, gamma 1, also known as PLCG1 and PLCgamma1, is a protein that in humans involved in cell growth, migration, apoptosis, and proliferation. It is encoded by the PLCG1 gene and is part of the PLC superfamily.

Fibroblast growth factor receptor 4 is a protein that in humans is encoded by the FGFR4 gene. FGFR4 has also been designated as CD334.

Fibroblast growth factor receptor substrate 2 is a protein that in humans is encoded by the FRS2 gene.

Fibroblast growth factor 5 is a protein that in humans is encoded by the FGF5 gene.

FGFR1 oncogene partner is a protein that in humans is encoded by the FGFR1OP gene.
Platelet-derived growth factor receptor A, also termed CD140a, is a receptor located on the surface of a wide range of cell types. The protein is encoded in the human by the PDGFRA gene. This receptor binds to certain isoforms of platelet-derived growth factors (PDGFs) and thereby becomes active in stimulating cell signaling pathways that elicit responses such as cellular growth and differentiation. The receptor is critical for the embryonic development of certain tissues and organs, and for their maintenance, particularly hematologic tissues, throughout life. Mutations in PDGFRA, are associated with an array of clinically significant neoplasms, notably ones of the clonal hypereosinophilia class of malignancies, as well as gastrointestinal stromal tumors (GISTs).

Autophosphorylation is a type of post-translational modification of proteins. It is generally defined as the phosphorylation of the kinase by itself. In eukaryotes, this process occurs by the addition of a phosphate group to serine, threonine or tyrosine residues within protein kinases, normally to regulate the catalytic activity. Autophosphorylation may occur when a kinases' own active site catalyzes the phosphorylation reaction, or when another kinase of the same type provides the active site that carries out the chemistry. The latter often occurs when kinase molecules dimerize. In general, the phosphate groups introduced are gamma phosphates from nucleoside triphosphates, most commonly ATP.
Fibroblast growth factor receptor oncogene partner 2 (FGFR1OP2) was identified in a study on myeloproliferative syndrome (EMS). The study aimed to identify the partner genes to the fibroblast growth factor receptor 1 (FGFR1) involved in the syndrome. Using the 5'-RACE PCR technique, FGFR1OP2 was identified as a novel gene with no known function.

Interleukin 17 receptor D is a protein that in humans is encoded by the IL17RD gene.
Clonal hypereosinophilia, also termed primary hypereosinophilia or clonal eosinophilia, is a grouping of hematological disorders all of which are characterized by the development and growth of a pre-malignant or malignant population of eosinophils, a type of white blood cell that occupies the bone marrow, blood, and other tissues. This population consists of a clone of eosinophils, i.e. a group of genetically identical eosinophils derived from a sufficiently mutated ancestor cell.

Pemigatinib, sold under the brand name Pemazyre, is an anti-cancer medication used for the treatment of bile duct cancer (cholangiocarcinoma). Pemigatinib works by blocking FGFR2 in tumor cells to prevent them from growing and spreading.